http://dx.doi.org/10.12795/spal.2007.i16.05
Escacena Carrasco, J. L. y García Rivero, D. (2007): “Selección sexual y éxito reproductivo en la protohistoria ibérica. Un enfoque evolucionista”, Spal 16: 69-92. DOI: https://dx.doi.org/10.12795/spal.2007.i16.05
Selección sexual y éxito reproductivo en la protohistoria ibérica. Un enfoque evolucionista
SEXUAL SELECTION AND REPRODUCTIVE SUCCESS IN IBERIAN PROTOHISTORY. AN EVOLUTIONARY APPROACH
José Luis Escacena Carrasco
Departamento de Prehistoria y Arqueología de la Universidad de Sevilla
Daniel García Rivero
Departamento de Prehistoria y Arqueología de la Universidad de Sevilla
Resumen: En relación con la Iberia protohistórica, los estudios acerca de la conducta sexual y de sus consecuencias sobre la reproducción de individuos y poblaciones son muy escasos; y prácticamente inexistentes los que han recibido un enfoque evolucionista. Este trabajo pretende analizar dicho tema desde la tendencia teórica de la Arqueología Evolutiva, es decir, desde una perspectiva darwinista.
Abstract: The studies about sexual behaviours and their consequences for the reproduction of individuals and groups in the Iberian Protohistory are few, and practically non existent those which have worked from an evolutionary perspective. This paper aspires to analyse these issues from the theoretical focus of Evolutionary Archaeology, that is, from a Darwinian perspective.
Palabras clave: Edad del Bronce, Edad del Hierro, sexualidad, reproducción, coevolución, genes, memes.
Key words: Bronze Age, Iron Age, sexuality, reproduction, coevolution, genes, memes.
PROBLEMAS DE TEORÍA, DE MÉTODO Y DEL REGISTRO
Fuera de los estudios sobre Paleolítico, la Teoría de la Evolución carece de aplicaciones en la tradición arqueológica hispana. Las bases epistemológicas de la Arqueología Evolutiva, línea en la que venimos trabando desde la Universidad de Sevilla, plantean que la fuerza primera que ha dado forma a la cultura humana es la selección natural según el sentido básico propuesto por Darwin. Por tanto, el enfoque aquí asumido pretende aplicar al comportamiento humano los principios científicos la Evolución por Selección. Desde sus comienzos en el siglo XIX, esta perspectiva ha tratado el tema de la sexualidad como uno de los principales aspectos en los que se hacen palpables las presiones selectivas. En ocasiones, incluso ha rechazado de forma explícita que otros cuerpos teóricos puedan decir algo al respecto, sea porque no les ha preocupado la cuestión, sea porque se ha calificado de “aberración” metodológica la extensión de su uso a tiempos prehistóricos[1]. Sin embargo, su aplicación a las sociedades antiguas ha tropezado con la escasez de datos que superen los puramente paleontológicos. Como tendremos ocasión de comprobar, tal carencia es sólo aparente. Una dificultad añadida proviene de que el conocimiento de las culturas protohistóricas debe de establecerse fundamentalmente a través del registro material que el comportamiento del hombre deja en el medio en el que habita, mientras que las investigaciones fundamentales en Teoría Evolutiva se han dirigido mayoritariamente hacia el componente somático humano.
Desde que Darwin diera a conocer su obra sobre cómo fueron apareciendo las distintas especies de organismos, el desenlace más coherente fue la aplicación de su teoría al nacimiento y evolución del hombre. Consecuente con su visión, él mismo publicaría a continuación su libro sobre los orígenes humanos. En él incluía además sus ideas relativas a la selección sexual (Darwin 1871), porque tal vez percibió muy pronto, como ahora se reconoce (Andersson 1994), lo perita que ésta resulta al analizar los costes y beneficios en la elección de pareja cuando hace que se hereden órganos adaptados expresamente para la función del cortejo, o incluso predisposiciones sensoriales a los estímulos de éste (Endler y Basolo 1998). Descartó así el propio Darwin que Homo supusiera una anomalía a su propuesta teórica, y trató el asunto, en consecuencia, desde el mismo enfoque metodológico propuesto previamente para los otros seres vivos. Sus consideraciones se extendían a las normas de conducta de los animales, hoy la etología. De esta forma, el darwinismo representó un marco epistémico creado para explicar tanto los caracteres somáticos y fisiológicos como las maneras de proceder de organismos y comunidades. No obstante, la visión monista darwiniana, que ve incapacitada a la selección natural para discriminar entre cuerpo y conducta con el resultado de diseñar el primero y no la segunda, ha sido normalmente rechazada incluso por quienes en los ambientes académicos se tienen por conspicuos evolucionistas[2]. Así, la arqueología ha sido definida como ciencia lamarckiana por el mero hecho de que la cultura humana se transmite por herencia aprendida (Querol 2001: 358), como si la imitación de comportamientos no fuera también propia de otros muchos animales. Asimismo, las tradiciones humanistas que abordan el análisis histórico, el antropológico o el sociológico, han proscrito las propuestas darwinistas cuando se aborda la evolución cultural de los grupos holocénicos, en el sentir quimérico de que la evolución habría concluido o de que el hombre sería ya dueño y señor de su destino. Sólo recientemente se han presentado trabajos de arqueología postpaleolítica bajo el enfoque evolutivo, algunos de los cuales cuentan con técnicas y procedimientos de análisis surgidos en la biología darwiniana. En esta tradición, las obras más significativas proceden tanto del ámbito anglosajón (Maschner 1996; O’Brien y Lyman 2000; Shennan 2002) como del habla hispana (Escacena 2000 y 2005; Martínez y Lanata 2002; García Rivero 2004; Muscio 2006).
El estudio de las sociedades protohistóricas de la Península Ibérica se ha abordado siempre desde alguna posición epistémica, nunca desde la inocencia teórica. Otra cosa es que se haya tenido consciencia de cuál era cada enfoque y que éste se haya explicitado a la hora de explicar los hechos. En cualquier caso, está extendida por doquier la paradoja por la que todas las perspectivas arqueológicas aceptan el darwinismo sin aplicarlo más que a la evolución somática. Esto ha edulcorado la teoría hasta hacerla aceptable por cualquier fe, sea ésta de índole religiosa o medre por los ámbitos académicos como programa político camuflado de cuerpo científico (Escacena 2002). Pocos investigadores han asumido la conclusión clave del darwinismo: que la causa generalizadora de los cambios es la selección natural. Someterse a dicho principio exige sustanciales mutaciones del pensamiento a la hora de aterrizar en los estudios antropológicos y arqueológicos. Por una parte, debe tratar a Homo sólo como un animal más en el conjunto de los seres vivos. Por otra, tendrá que asumir que la conducta humana de hoy es, tanto como la de ayer, una mera manifestación evolutiva. Por lo cual, la mayor parte de nuestro comportamiento es, como quien dice, «ruido de fondo», en el sentido de que escasean notas singulares relacionables sólo con la especie a la que pertenecemos.
Por diversas causas, la práctica arqueológica ha considerado en extremo difícil la aplicación del pensamiento darwiniano. Como acabamos de adelantar, la razón más poderosa para esta renuncia ha sido tal vez la dificultad para reconocer la naturaleza animal humana. Diversas filosofías y religiones se han encargado siempre de lo contrario, es decir, de hacernos creer que somos una clase de rex mundi. Tamaño antropocentrismo ha sido desde luego un pensamiento muy adaptativo, por lo que la propia selección natural se ha afanado en extender sus armas filosóficas, políticas y religiosas. De esta forma, hemos explotado un amplio espectro de ecosistemas sin problemas de conciencia. O pretendemos ahora manejarlos supuestamente mejor desde la mentalidad ecologista, tan antropocéntrica en el fondo como otras muchas creencias de índole religiosa. Es así como hemos incrementado históricamente la aptitud de individuos y poblaciones, lo que ha contribuido sobremanera a nuestra propia reproducción. Entre los prehistoriadores y arqueólogos hispanos, pero no sólo entre ellos, escasean los consecuentes con los planteamientos de la evolución por selección. De ahí nace una práctica arqueológica que participa de un número cada vez mayor de tendencias teóricas. Todas ellas, en cualquier caso, únicamente han asumido a un Darwin demediado, útil para explicar los cambios corporales y fisiológicos pero inapropiado según esos otros planteamientos para hacer lo mismo con la conducta.
Las lecturas no darwinistas de la Historia han visto en Homo al ingeniero de su propio comportamiento. Desde esas posiciones, nuestra especie sería autora de un ingente número de revoluciones, avances tecnológicos y normas morales pensadas desde lúcidas mentes para el progreso individual y colectivo a corto, medio y largo plazo. Por todas partes surgen quienes ven en nuestro devenir histórico una suerte de magisterio para la vida futura. Es más, la misma disciplina arqueológica se ofrece como una cuasi redención de las injusticias y desigualdades humanas, porque algunas de sus tendencias teóricas prometen ese futuro idílico. Nuestro enfoque evolucionista parte de asumir, empero, que las leyes que nos conducen por el mundo han sido y son forjadas por un conjunto de fuerzas no controladas por la voluntad personal ni perfiladas para su funcionamiento a largo plazo por ninguna mano ingeniera, ni eugenésica ni eumemésica[3]. Como afirma el dicho popular “el hombre propone y Dios dispone”, la conducta humana experimenta constantemente variación, pero los «inventos» que origina no llegan a generalizarse sino por la fuerza selectiva de causas no dominadas por la sociedad ni por sus individuos. Con referencia al ámbito concreto de la sexualidad y de sus consecuencias sobre la demografía, tampoco es el hombre, en consecuencia, el responsable del supuesto proyecto teleológico de las normas que han regido y rigen nuestra conducta. Para un enfoque evolutivo, los tabúes sobre el sexo son tan naturales como la feromona de una mariposa, y ambos contribuyen al incremento poblacional de las especies que los portan. En consecuencia, es lícito analizar con el mismo paradigma científico tanto las estrategias humanas de cortejo como los nidos de las golondrinas o las elucubraciones teológicas más alambicadas.
Las teorías que han entrado al estudio de la Prehistoria Reciente y Protohistoria hispanas han asumido casi siempre visiones finalistas de la evolución humana. Nuestras bibliotecas rebosan de análisis enmarcados en el Historicismo Cultural. Menos común fue hasta hace poco en las universidades españolas el enfoque marxista, y todavía más escasas las perspectivas epistemológicas conocidas como Arqueología Procesual y como Materialismo Cultural. Desde muchas de estas lentes, la historia humana se ha interpretado como camino de progreso, y se ha identificado por tanto «evolución» con «mejora». Este tema ha sido tratado en profundidad por la Teoría de la Evolución al menos desde Darwin, y sólo ha llegado al resultado de aceptar, en cualquier caso, que la evolución sí conduce de hecho al aumento de la complejidad, pero sólo porque ésta, entendida como diferenciación de las partes de un todo, origina cada vez más nichos ecológicos y soporta por ello más vida. Aunque la curva que representa esta tendencia puede contener dientes de sierra, en su dirección general no escapa a esta norma el devenir histórico de la Península Ibérica, desde los tiempos paleolíticos hasta hoy.
Pero confesar que las poblaciones protohistóricas hispanas experimentaron tal incremento en su complejidad interna no presupone aceptar que la evolución de los grupos humanos locales se limitó a un desarrollo lineal sin quiebra. La velocidad de la evolución ha conocido dentro de la propia escuela darwinista sustanciosos cambios, que van desde el gradualismo sospechado por Darwin hasta la propuesta saltacionista del “equilibrio puntuado” de N. Eldredge y S.J. Gould (1972: 82-115). Casi ningún investigador de la Prehistoria Reciente y Protohistoria ibéricas ha intuido siquiera que tales asuntos sobre el ritmo evolutivo vayan con su parcela de trabajo. Tampoco han dado el visto bueno los especialistas, sobre todo desde los años sesenta del siglo XX para acá, a rupturas significativas en la secuencia de transformaciones que conduce desde las primeras comunidades campesinas, las neolíticas, hasta las de la Edad del Hierro. Frente a estas posiciones, tomar conciencia del ser exclusivamente animal de Homo acarrea de forma ineludible la aceptación de eventos traumáticos diversos que pudieron romper una teórica cadena evolutiva directa. Tales episodios estarían en condiciones de acelerar el cambio cultural, no tanto por la novedad que podían suponer como por el aumento de la diversidad que provocaban, base para la posterior labor de criba de la selección natural. Esto pudo llegar al extremo de no hacer reconocible la paternidad de ciertas tradiciones en las poblaciones precedentes dentro de la misma región. Otras veces, esos colapsos habrían originado despoblamientos en diversas épocas y ambientes geográficos. En estos aspectos, el pensamiento no darwinista, consecuente con la idea de que el hombre es la cima de la creación y el dueño de la naturaleza y de su propio destino, lo ha creído capaz de solventar cualquier problema ecológico. Así, y para que valga como ejemplo bajoandaluz para la época protohistórica que aquí estudiamos, una y otra vez la falta de documentación se rechaza como síntoma de vacío demográfico, y se hecha mano para explicarla de dos argumentos: la carencia de investigaciones (cf. Ferrer y De la Bandera 2007: 49) y la existencia de comunidades que no habrían dejado huella arqueológica debido a su vida itinerante (cf. Ruiz-Gálvez 1995: 161-162). Tal renuncia a asumir hiatos en la secuencia de ocupación antrópica de algunos espacios ibéricos acarrea irremediablemente visiones evolucionistas lineales difíciles de compaginar con los eventos saltacionistas aprobados hoy por el darwinismo.
La Protohistoria de la Península Ibérica fue, por el contrario, un hervidero de heterogeneidad. Es imposible entenderla como un todo. Fue diversa en sus ingredientes culturales y en las fuentes de las que éstos surgieron, tan diversa como aparece en el registro paleolingüístico la situación que precedió a la conquista del territorio por Roma. En este sentido, J. De Hoz (1991: 40) ha propuesto un primer milenio a.C. similar a un mosaico de grupúsculos cuya movilidad explicaría la extrema complejidad toponímica. De la misma forma, nuestra concepción de este tramo histórico nos impide tratarlo de forma global y homogénea. De ahí que nos limitemos a proponer diversos caminos de entrada al conocimiento de los grupos humanos de entonces en los temas que dan cuenta de la sexualidad y la reproducción. Ello no implica que, demostrada tal o cual característica para un momento determinado, sea posible llevar de forma automática esa explicación a tiempos anteriores o posteriores cuantos siglos o milenios se nos antojen.
SOBRE REPLICADORES
Quienes han asumido la propuesta darwinista, especialmente a la hora de ofrecer visiones monistas del ser humano, sostienen que la evolución ha transcurrido, en su nivel básico, entre unidades de competencia: los replicadores biológicos. En la herencia corporal, los códigos que transmiten a la descendencia las características somáticas estarían alojados en el ADN. Los genes constituirían así los replicadores elementales, y en ellos se produciría, como quieren los neodarwinistas, el principal nivel de selección. Según esta tendencia, en los genes estarían contenidas las directrices esenciales que tutelan también la conducta. Instalados en el interior del núcleo celular, han sido reivindicados por estudios posteriores a Darwin, en especial desde que las inferencias mendelianas se acoplaron a la teoría evolucionista a partir de la Síntesis (Wilson 1980). Como parece que Darwin no tuvo acceso a los descubrimientos de Mendel, consideró que el plano en que actuaba la selección lo componían, por este orden de importancia, el individuo y el grupo. Luego, ciertos especialistas han considerado, en atención a la propuesta de S. Wright (1945), que el filtro selectivo podría trabajar entre poblaciones o conjuntos de individuos mucho más de lo sospechado inicialmente por el padre de la teoría. Dicha modalidad, que se ha denominado selección de grupo, ha sido una constante fuente de sorpresas a la hora de comprender la evolución, sobre todo cuando se han querido explicar determinadas características de las simbiosis mutualistas en general y de las especies gregarias en particular. Por eso representará dentro de nuestro enfoque un efectivo instrumento de trabajo.
Se ignora aún hasta qué grado de precisión el genotipo diseña los comportamientos humanos. Los sociobiólogos han sido propensos a aceptar que la conducta viene fuertemente determinada por la herencia genética. En cambio, quienes rechazan tanto poder para el legado del ADN asignan al aprendizaje el papel esencial en la labor de perfilar los comportamientos concretos, especialmente aquellos que no son comunes a toda nuestra especie. En cualquier caso, todos se muestran de acuerdo en que los detalles singulares de cada grupo sólo pueden sustentarse en la existencia previa de condiciones comunes que sí estarían controladas por los genes. Esas órdenes primarias supondrían no más que la condición necesaria, no la suficiente.
El darwinismo ha trabajado mucho menos en la transmisión cultural que en la herencia somática y fisiológica, y menos incluso que en el estudio de la conducta innata. Es bastante reciente, de hecho, el uso en este marco teórico de términos universalmente admitidos para referirse a un mismo fenómeno, característica u observación, requisito casi imprescindible para trabajar con corrección científica. Desde 1976 está disponible la voz meme con el significado de unidad mínima de transferencia cultural (Dawkins 1979 [1976]: 277-293). No obstante, debemos a S. Blackmore (2000) el mayor desarrollo de este concepto. Frente a los genes, responsables del legado somático y de la actuación innata, los memes se encargarían de replicar las ideas y la conducta compleja, transferidas ahora mediante la imitación. Se desplazan estos últimos diacrónicamente para llevar a cabo la transmisión vertical, desde los progenitores a la descendencia, de la conducta aprendida; pero cuentan con más facilidad que los genes para circular en horizontal en un tiempo dado. De esta segunda forma, serían responsables de las interacciones culturales entre partes sincrónicas, sean individuos o poblaciones. Si el genotipo consiente de hecho alguna variación mediante la plasticidad fenotípica, la transmisión memética muestra mayor elasticidad. Esto no supone que los cambios escapen a la labor de criba de la selección natural.
El encargo de genes y memes es originar homólogos a sus ascendientes. No obstante, en los dos casos los caracteres de la progenie son copias «erróneas» de los paternos, porque nunca crean réplicas idénticas de los mismos. Mutaciones génicas y recombinación cromosómica por una parte, e «infidelidades» culturales por otra, originan heterogeneidad por doquier, las primeras en el plano somático y las segundas en la conducta aprendida. Esta diversidad es el nicho ecológico de la selección. Sin variación no existe evolución, simplemente porque la selección natural no puede optar entre expectativas diversas. En este contexto, los propios darwinistas discrepan acerca de si los genes controlan a los memes o son estos últimos los que han tomado las riendas evolutivas. Aunque esta diatriba ha tomado cuerpo en algunas publicaciones (p.e. Alexander 1994: 74; Blackmore 2000: 143-177), tal vez estemos de nuevo ante una caída inconsciente en visiones no monistas del hombre. De ahí que no pueda extrañar una simbiosis mutualista entre memes y genes, a la manera de cualquier alianza biológica en la que todas las partes de un todo salen beneficiadas a través de un proceso coevolutivo engranado. Es ese pacto el que ha llevado a la especie Homo sapiens a su dispersión por todos los ecosistemas terrestres, y hoy ya también por algunos extraterrestres.
En cuando al tema de nuestro artículo, genes y memes han cooperado a lo largo del tiempo para dotar a la selección natural de la materia prima que define la sexualidad humana. Por tanto, para la gente que habitó la Iberia protohistórica, como para las culturas actuales, las costumbres relativas al sexo y sus efectos sobre la reproducción determinan una herencia biológica difícilmente evitable. Igual que en otras muchas expresiones de nuestro comportamiento, aquí se manifiesta también con toda su fuerza el «ruido de fondo», es decir, un proceder que hace de nuestras pautas sexuales y reproductivas un legado común a múltiples especies[4].
SELECCIÓN SEXUAL
La lectura de Malthus persuadió a Charles Darwin del importante papel de la reproducción diferencial para comprender cómo se generalizan o desaparecen los caracteres heredados. Como una simple operación aritmética de suma y sigue, la cuantía de descendencia es directamente proporcional a las posibilidades de que las características de ésta se prodiguen en las siguientes camadas. Tal fenómeno origina procesos de retroalimentación positiva que acaban generalizando mutaciones a amplios sectores de la población. Desde tal perspectiva, el principio evolucionista de «supervivencia de los más fuertes» fue tomando el sentido de «supervivencia de los más aptos», entendidos estos últimos como los que, al prolongar su vida, potencian las probabilidades de engendrar descendencia.
Desde un enfoque darwinista, nuestra conducta sexual es el resultado de un largo devenir evolutivo en el que han triunfado las costumbres de quienes más se han reproducido. Pero el concepto de reproducción debe verse en este marco como crecimiento neto de la población. De hecho, no siempre el aumento demográfico de un grupo va unido a su mayor índice de natalidad. Así, la aparición de la guerra ocasionó mayor éxito reproductivo en las comunidades que la aplicaron con más efectividad, con lo que quedaba garantizada la propagación de sus genes y de sus memes (incluido en este último lote el propio recurso a la violencia colectiva organizada). En este terreno, nuestro parentesco parece estar más cerca ahora de los agresivos chimpancés que de los apacibles bonobos.
CARIÑO Y AGRESIVIDAD. ESTRATEGIAS Y COMPETENCIA EN LA ELECCIÓN DE PAREJA
Puede parecer paradójico que las emociones amorosas surgieran a lo largo de la evolución de los animales a partir de los sentimientos agresivos o de forma paralela a ellos. La reproducción sexual nació después de otras fórmulas de replicación. Es anterior la clonación, pero también en algunos casos la partenogénesis, procedimientos que consiguen duplicados idénticos a los progenitores. La agresividad es efectiva en contextos en los que la pitanza escasea, se dé esto por disminución de la oferta o por aumento de la demanda. Nació pronto en la historia de la vida, parte de la cual se organiza y medra a costa del sacrificio de otros seres vivos. Por el contrario, los gestos de amor y el altruismo tuvieron más posibilidades de manifestarse cuando surgió el sexo o conforme aumentaba el cuidado de la prole. No extraña por tanto que las relaciones amorosas y las atenciones parentales hacia las crías hayan coevolucionado a través de acciones y características muy parecidas. Cuando los recursos son exiguos o están irregularmente repartidos en tiempo y espacio, el amor hacia lo propio conduce en ocasiones a la agresión hacia lo ajeno. La protección de la familia conlleva con frecuencia la apropiación de un territorio o de un nicho ecológico, una apetencia que también expresan otros individuos o poblaciones cercanas. El conflicto y el afecto se muestran así como dos caras de una misma moneda.
En muchas culturas del mundo antiguo fueron casi unidas masculinidad y belicosidad. Tanto se asoció una a la otra, que fue una norma femenina valorar en el hombre, sobre todo cuando se trataba de elegir pareja, aquellos aspectos que realzaban el valor, la destreza en el combate, el honor militar, el grado que cada varón ocupaba dentro del grupo de guerreros, etc. En relación con hipotéticos problemas futuros –y siempre pensando en términos evolutivos no necesariamente conscientes para los protagonistas– la hembra calibraba así las posibilidades de protección de la prole a la vez que las garantías en el suministro de recursos para la familia. Son los memes transferidos culturalmente los encargados de fomentar en la mujer el ideal de compañero. La elección de un modelo distinto que acarreara menos beneficios reproductivos al hogar estaría proscrita por la selección natural, o al menos se perpetuaría en proporción mucho menor. Cualquier pauta cultural que condujera a una hembra a la elección errónea, estaría seriamente en peligro de supervivencia por el mero hecho del propio fracaso reproductivo parejo a la decisión equivocada.
Si son correctos tales pronósticos darwinistas, estaríamos en condiciones de encontrar huellas arqueológicas relativamente claras de los mismos en la Protohistoria hispana. El guerrero valeroso quedó por supuesto representado en diversos tipos de iconos, ya en exvotos ofrecidos a los dioses ya en grabados sobre estelas de piedra (fig. 1). Si una de sus características consiste en realzar la fortaleza y la robustez a través de adornos corporales, los tocados de plumas que portan los arqueros del arte neolítico levantino supondrían ya un precedente prehistórico de esta preocupación por reforzar la masculinidad. Esos «adornos» enfatizan la musculatura de los brazos y de las piernas, con especial incidencia en este segundo caso en las pantorrillas. Dicha norma está registrada en diversas comunidades primitivas, pero es también una regla en los uniformes de los ejércitos históricos, que tienden a acrecentar la anchura del tórax. El acceso al cuerpo de soldados delimita con frecuencia en muchas comunidades humanas los ritos de tránsito que se celebran en el paso a la mayoría de edad de los muchachos, en una identificación entre la plenitud sexual y la integración plena del joven en el conjunto de guerreros de la comunidad. Entre los íberos y celtíberos los varones llevaban a cabo ejercicios atléticos parecidos a las acrobacias y carreras plasmadas en el arte naturalista neolítico del Levante español. Tales manifestaciones de destreza física y de gestos marciales se hacían en presencia de las mujeres, quienes podían así contrastar las virtudes de los hombres casaderos (Caro Baroja 1946: 157-175). Es posible que esta costumbre hispana protohistórica provenga de poblaciones mucho más antiguas asentadas en la zona oriental de la Península Ibérica al menos desde el cuarto milenio a.C. Así, por su capacidad para elegir, la mujer podría haber contado con un papel más activo del que en principio podría atribuírsele; y tal situación le confería un control efectivo sobre los miembros más próximos de su vida familiar, circunstancia reconocida cuando se han comparado los distintos ámbitos en los que se manifiesta tradicionalmente el poder: el público o político, masculino, y el emocional, femenino (Hernando 2007: 170-171).
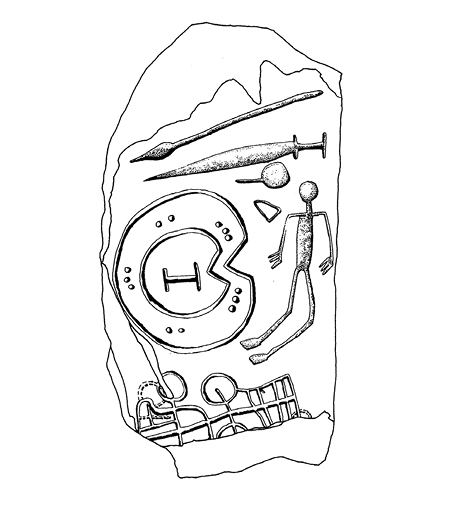
Figura 1. Guerreo con su panoplia representado en la estela de Solana de Cabañas.
Que la masculinidad va en estas culturas antiguas unida al valor del guerrero quedaría demostrado hasta la saciedad en la pléyade de exvotos prerromanos en los que se representan varones desnudos armados, uno de cuyos principales atributos es precisamente el pene erecto. No puede extrañar entonces que las mujeres llevaran a los santuarios tales figurillas como agradecimiento a las divinidades por haber logrado el compañero ideal (fig. 2). Entendemos por tanto, al contrario de lo que se hace normalmente (p.e. Rueda 2007), que para conocer a la mujer protohistórica podrían valer más las representaciones masculinas que las de ellas mismas, y viceversa.

Figura 2. Ideal de varón para la mujer íbera. Exvoto en bronce (según Nicolini 1977).
La fuerza y el vigor atribuidos a la masculinidad se han vinculado en muchas culturas a los animales más fieros, potentes y temidos de los distintos ecosistemas en que dichas sociedades han vivido. En las culturas de la Protohistoria hispana los bóvidos tuvieron ese significado. Por eso muchos grupos vieron en su imagen el símbolo esencial de dichos atributos; y en una parábola simplificadora que puede considerarse una indudable sinécdoque, el toro omnipotente acabó reducido a su cornamenta. Desde esta asimilación de la parte al todo, la asociación más clara entre el valor del guerrero y la potencia del toro –potencia referida posiblemente tanto a su fuerza como a su ímpetu sexual– se manifiesta en las estelas grabadas de finales de la Edad del Bronce, cuya mayor expansión implica al cuadrante suroccidental de la Península Ibérica. Y si la pujanza viril y la fuerza del toro se alojan simbólicamente en sus cuernos, a mayor tamaño de éstos mayor abundancia de aquellos caracteres. No sería aleatorio, en consecuencia, que la representación de los jefes plasmada en esos monolitos exhiba a veces una acusada desproporción entre el tamaño del personaje y el de las astas que rematan el casco (fig. 3).
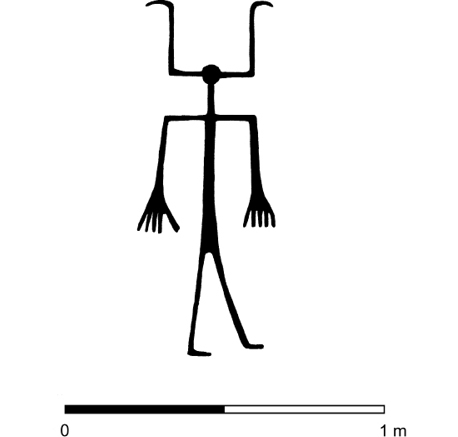
Figura 3. Figura de guerrero de la estela de El Coronil (a partir de Izquierdo y López Jurado 1998).
A raíz de la colonización de griegos y fenicios se introducen en el extremo Occidente unas creencias relacionadas con el toro en las que este significado se hace aún más explícito. Los bóvidos machos, y desde luego nunca castrados, fueron la ofrenda predilecta del Baal cananeo, mientras que las novillas se sacrificaban a Astarté. La liturgia ugarítica plasmada en los textos referidos al ciclo baálico constituye el precedente remoto de estos mitos que a comienzos del primer milenio a.C. llegan hasta el Mediterráneo oeste y hasta el Atlántico. Esas referencias escritas dan buena cuenta de estos vínculos, y comparan constantemente al dios con el toro y con los atributos sexuales que lo hacen macho. Paralelamente, la diosa es asimilada a la vaca-esposa del toro y se le dota de los caracteres propios de la mujer madre. La alusión a Baal y la llegada de su culto a la Península Ibérica al final de los tiempos prehistóricos proporcionan un abundante terreno en el que trabajar, y nos servirán para tratar un último aspecto dentro aún de este apartado.
A comienzos del primer milenio a.C., Baal es en efecto el Señor de los fenicios, adorado bajo distintas advocaciones locales que se dispersan por las colonias conforme avanza hacia poniente la colonización cananea. Es Señor como tratamiento de respeto –eso significa su propio nombre–. Pero nada nos extrañaría que, en los momentos de la diáspora fenicia por Occidente, dicho apelativo tuviera ya un sentido semántico similar a una de las acepciones del término latino senior (viejo), con las correspondientes connotaciones de respetabilidad que la ancianidad tuvo para las culturas antiguas mediterráneas. En tal caso recibiría cabal explicación la existencia de figurillas sagradas en las que un dios (¿Melqart como Señor de la ciudad?) se materializa en la imagen de un anciano de larga y poblada barba (fig. 4). Entonces, esta cualidad se pudo calificar más como algo meritorio que como señal de caducidad. Por eso no puede soslayarse que la situación acomodada que el varón adulto solía lograr fuera otra razón bien vista por las mujeres de la época a la hora de optar por un pretendiente. Si el joven guerrero garantizaba la defensa grupal por su fuerza y valor, el varón entrado en años suponía el bienestar de la familia por su posición consolidada en la estructura socioeconómica de la comunidad. Desde esta premisa, podrían tener explicación los diferentes gustos manifestados respectivamente por dos mujeres en la fábula que Viriato narró a los habitantes de Itucci (Martos, en la provincia de Jaén)[5], con la que J.C. Bermejo ha defendido la existencia probable en el mediodía ibérico de lo que podría denominarse el “mito de la calvicie de la luna”. El cuento, transmitido por Diodoro Sículo (XXXIII, 7,5), estaría aludiendo tal vez a la existencia de tradiciones religiosas en las que dos rivales femeninas competirían por alcanzar los favores amorosos de la luna, que aparece en la leyenda como personaje masculino. A una de las contendientes le agradan los pelos oscuros de éste (imagen del joven), con lo que decide depilar sus canas; la otra mujer hace lo contrario por preferir los cabellos blancos (imagen del viejo), hasta que entre las dos dejan al novio mondo y lirondo (Bermejo 1984: 181-191; 1986: 45-46). Esta doble estrategia mostraría la coexistencia en aquella sociedad de opciones diversas a la hora de elegir macho por parte de las hembras. Caro Baroja (1976: 187-188) leyó todavía más allá en esta historieta: quiso ver en ella la posible existencia en la Lusitania, tierra de origen de Viriato, de matrimonios polígamos. En cualquier caso, frente a esta doble posibilidad de elección por parte de las mujeres, los hombres han optado siempre por mujeres jóvenes, lo que se explicaría desde un análisis evolucionista por el fracaso reproductivo para el varón si hubiese preferido una compañera anciana (Jones 1996).

Figura 4. Exvoto en bronce que representa a un dios fenicio ya maduro. Procede de los alrededores del santuario gaditano de Melqart en Sancti Petri.
COEVOLUCIÓN ENTRE GENES Y MEMES SEXUALES Y DE LA CRIANZA
Los gestos específicos de las relaciones sexuales, en especial los de seducción y cortejo que preceden a la cópula, muestran en muchos animales un asombroso parecido formal con los que caracterizan a la relaciones entre los padres y sus crías. La descendencia de muchas especies publicita su propia existencia con sonidos, con colores llamativos o con movimientos que reclaman la asistencia de los progenitores. Los biólogos saben que, en los polluelos de muchas aves, hay una razón directamente proporcional entre la apertura de la boca y el logro de comida en el nido. Funciona la misma ley cuando la prole cuenta con reclamos visuales. Pedir sustento se acompaña en algunos casos de gestos concretos cuya misión es obtener lo más posible, lo que establece una competencia directa con el resto de la camada. Abundan también los casos en que los guiños amorosos entre sexos opuestos muestran un repertorio conductual parecido a esas tácticas pueriles. Desde tal perspectiva, podría afirmarse que la selección natural ha propiciado cierta homogeneidad en los signos destinados a demandar atención, sean éstos los de las relaciones paterno-filiales sean los que se dan en la pareja macho-hembra. De esa forma, podría predecirse que el cortejo entre humanos, es decir, las acciones más o menos ritualizadas por las que una parte acapara la atención de la otra y minimiza la competencia intrasexual, cuente también con fenómenos similares de neotenia. El trabajo en esta hipótesis ha llegado a reconocer actitudes simbólicas y normas de conducta que cumplen efectivamente dicha constante (cf. Eibl-Eibesfeldt 1995: 130-155). El galanteo estaría destinado, pues, en hombres y mujeres a merecer la atención del opuesto preferido. En este ambiente competitivo, habrían salido adelante las estrategias que coincidieron en parecido externo con actitudes de paidomorfismo, esto es, las que semejaban formalmente las acciones que los bebés llevan a cabo para solicitar afecto y amor por parte de sus mayores.
El rostro de los niños lactantes tiene diversas características cromáticas, todas ellas atractivas para los padres. Una es el enrojecimiento de los labios, otra el de los carrillos. Unido a tales rasgos infantiles, un tercer elemento característico es el mayor tamaño del cráneo, en relación con el del resto del cuerpo, que el mostrado por los adultos. Un cuarto elemento añadido es una notable escasez de pelo en la cabeza. Dichas peculiaridades acentúan en los mayores cierta propensión innata a mostrar afecto a los pequeños y a cubrirlos de lisonjas y atenciones. Cuando practicamos el sexo, las reacciones químicas que preceden y acompañan al clímax estimulan cierto sofoco en los adultos (Morris 1995: 60 y ss.), acompañado de una ruborización similar al pavo espontáneo del lactante. Una interpretación no darwinista de la evolución podría defender que las modas de embellecimiento corporal habrían tendido a imitar esas situaciones, fuera de manera voluntaria y consciente o no. Sin embargo, dar preponderancia a la selección natural lleva a considerar la llegada a esta semejanza por un proceso de convergencia. Así, surgida en algún momento y lugar, fuera cual fuera la causa, la costumbre de adornar el rostro con coloretes, dicha pauta conductual habría aprovechado en su propio beneficio evolutivo –entiéndase a favor de su expansión– la propensión humana a que los adultos sientan simpatía por la imagen infantil. Por consiguiente, una lectura darwiniana defendería que se trata de una similitud formal analógica[6]. Los polvos rosados en los pómulos, usados ya en muchas culturas mediterráneas antiguas para el arreglo femenino, suponen un reclamo óptico hacia el varón. Desde un análisis evolucionista representarían gestos meméticos que la selección natural, manifestada en este caso bajo la apariencia de selección sexual, ha hecho confluir con los genotipos que producen un fenotipo similar en las crías, con el resultado en este segundo caso de lograr atenciones parentales. Parecido efecto tiene el hábito de enrojecer los labios. En este caso, dicha costumbre centra en la boca los gestos de cuidado y de atención amorosa, como hace cualquier adulto cuando proporciona comida a los bebés. La norma de pintarse de rojo se inició al parecer en tiempos prehistóricos muy remotos. En su origen pudieron estar implicados impulsos rituales de signo religioso o simbólico, porque ese color, rociado sobre enfermos o cadáveres, podría ejemplificar el refuerzo de la salud y hasta la donación de una nueva vida por su parecido cromático con la sangre. En cualquier caso, sea o no esto lo que puso en marcha el uso, a nuestra explicación le interesan menos las causas que los resultados. De hecho, muchos darwinistas reconocemos que, ante la imposibilidad de averiguar en múltiples ocasiones las voluntades con que los fenómenos históricos se inician, la meta genuina de la investigación evolucionista debería atender sobre todo a las razones por las que las costumbres se expanden y generalizan, es decir, a las causas de su replicación exitosa (cf. Rindos 1990: 3-9).
La arqueología protohistórica hispana cuenta con evidencias claras del uso de cosméticos. Los datos más explícitos proceden del mundo de los íberos, donde la mujer se representa con los labios pintados en la estatuaria que en mejores condiciones ha llegado hasta nosotros. La Dama de Baza constituye uno de esos testimonios (fig. 5), porque sus labios se tiñeron de un intenso rosa (Presedo 1973: 42). Pero no faltan ejemplos en rostros femeninos plasmados sobre cerámica. Así, los cachetes coloreados caracterizan a la cara del denominado Vaso de la Pepona (fig. 6). Son quizá testimonios mucho más viejos los frascos prehistóricos repletos de almagra (cf. Martí y Hernández 1988: 27; Acosta y Pellicer 1990: 103 y fig. 8, nº 20). No obstante, no deja de presentar dificultades relacionar estos ejemplos neolíticos con el embellecimiento de la mujer, porque otras explicaciones posibles hablarían de rituales religiosos achacables tanto a varones como a mujeres.

Figura 5. Rostro de la Dama de Baza (según Presedo 1973)

Figura 6. Cara femenina con coloretes en los pómulos (según Pericot 1979). En la barbilla muestra lo que parece un tatuaje facial.
Cuando se comparan con los ojos de los adultos, los de los bebés son mayores en proporción al tamaño de la cara. Estas facciones estimulan igualmente la atención de los padres y parientes del recién nacido; también la de las personas mayores que lo observan. Por eso, si nuestra propuesta tiene visos de verosimilitud, pintarse el contorno de los ojos para agrandarlos representaría de nuevo una coincidencia casual con la fisonomía infantil, pero ahora como táctica para seducir al varón por parte de las mujeres. Tampoco esta práctica puede catalogarse, pues, como homología evolutiva, porque la característica física de los ojos infantiles se hereda por los genes y la costumbre cosmética por los memes, es decir, por lo que la antropología cultural ha denominado comúnmente enculturación. En la arqueología protohistórica hispana contamos con las grandes pestañas de bronce que formaban parte de un posible rostrillo funerario de la mujer inhumada en el sarcófago fenicio femenino de Cádiz (Blanco y Corzo 1981: lám. 21, d) (fig. 7). También estaban pintados los ojos, las cejas y las pestañas de la Dama de Baza, aunque es posible que algunos de esos detalles, en especial los que aluden al vello, no indiquen necesariamente una pintura real en la mujer allí representada, sino que constituyan un recurso final del escultor para marcar esos elementos anatómicos, de la misma forma que coloreó de rosa también las manos sólo con la intención de sugerir el tono natural de la piel femenina. El comienzo del hábito de pintarse los ojos nos preocupa poco desde la teoría evolutiva, sobre todo porque cualquier hipótesis imaginada podría quedarse en pura fantasía ante la imposibilidad de someterla al principio popperiano de la falsación. En cualquier caso, se conocen algunas razones de por qué lo hicieron los egipcios. En el país del Nilo se inició al parecer como uso terapéutico, porque los ungüentos en los párpados y sus alrededores permitían retener el polvo del desierto antes de su entrada en el ojo, a la vez que filtraban la potente luz diurna de la zona. Por tal motivo, la pintura de ojos se usó allí por los dos sexos, de donde se sigue que su posterior reserva al exorno femenino en muchas culturas mediterráneas pudo evolucionar en paralelo al valor estético con que hoy cuenta tal moda. En cualquier caso, al teñir los párpados con colores oscuros se consigue el efecto óptico de agrandar los ojos y de hacer más blanco el vítreo, todo ello por el fenómeno físico denominado “contraste de brillos simultáneos” (Purves y otros 2003: 58) (fig. 8).

Figura 7. Pestañas de bronce del ajuar funerario del sarcófago fenicio femenino de Cádiz.

Figura 8. Contraste de brillos simultáneos. El color de los círculos centrales es exactamente el mismo en ambos casos, aunque parece más claro el de contorno oscuro.
El embellecimiento de la mujer ha confluido también con la anatomía de los niños en un carácter de mucha personalidad: la proporción de tamaño entre el cráneo y el esqueleto postcraneal. Las dimensiones de la cabeza son, en relación con las del resto del cuerpo, siempre más grandes en los pequeños que en los adultos. Tanto atraen estos rasgos paidomórficos a los mayores, que la publicidad los explota con insistencia. Así, para atraerse los favores de quienes los contemplan, los muñecos para juegos han mimetizado a la infancia en algunos de sus rasgos. Es paradigmática en este sentido la evolución seguida por el aspecto externo de Mickey Mouse, que pasó de representar a un viejo múrido cascarrabias a la imagen de un gracioso, aniñado y gentil ratoncillo (Gould 1994: 83-94). Podría verse aquí la causa de la distorsión intencionada del cráneo que se llevaba a cabo en algunas culturas antiguas, lograda con la aplicación de fuertes ataduras desde muy corta edad para lograr un desarrollo dirigido de los huesos que envuelven el encéfalo. En cualquier caso, no se conocen esqueletos protohistóricos hispanos ni representaciones artísticas con tales deformaciones óseas. Pero, como esta costumbre podría acarrear peligros para la salud, un efecto similar y menos dañino podía conseguirse con la adecuada manipulación del pelo. De tal guisa, los peinados y aderezos crinales, y hasta los postizos usados para dar volumen a la cabellera, podrían revelarse como dispositivos culturales que aprovechaban la atracción innata que en los adultos produce esa especial característica infantil. Por enésima vez, memes y genes se expresarían en una simbiosis mutualista en la que las dos partes de la relación habrían tirado desde el mismo foco selectivo. Aparece aquí de nuevo la figura de la convergencia evolutiva en forma de parecido analógico. Ejemplo hispano de dicho fenómeno podrían ser los múltiples perifollos con que las mujeres prerromanas tocaban sus cabezas. Recogidos ampliamente en la escultura en piedra de entonces, por ejemplo en la Dama de Elche (fig. 9), contamos no obstante con modelos aún más arcaicos en las «estelas diademadas» protohistóricas, que aluden a personajes femeninos con desproporcionadas cabezas y ricos atavíos (Celestino 2001: 254-260) (fig. 10). Éstas resultan especialmente grandes si se comparan con las de sus compañeros masculinos (fig. 11). Estrabón contó además que las mujeres de ciertas tribus hispanas se rapaban en parte la cabeza, lo que podría leerse de nuevo como una afinidad analógica con características infantiles por la ya citada escasez de vello típica de los bebés. En cualquier caso, la numerosa iconografía femenina en la que aparece la norma contraria, es decir, el intento de acrecentar la cabeza, sugeriría que esta otra moda medró en la Protohistoria hispana sólo como conato de parecido neoténico no primado por la selección natural frente a otras estrategias rivales (fig. 12). He aquí el texto más significativo de Estrabón (III, 4, 17) que refiere la costumbre:
Una cosa bárbara parece ser también el adorno de ciertas mujeres que describe Artemidoro. Según él en algunas regiones las mujeres llevan collares de hierro con unos cuervos (ganchos en forma de pico de cuervo) doblados sobre la cabeza y extendidos bastante por delante de la frente. Y sobre estos «cuervos» cuando les gusta ponen el velo, de manera que, extendido, da sombra a la cara, y esto les parece ser un adorno. En otros sitios ponen sobre el cogote una pequeña pandereta redonda que cubre la cabeza hasta las orejas aumentando poco a poco de altura y anchura. Otras mujeres se quitan el pelo de la parte anterior de la cabeza tanto que ésta luce (aún) más que la frente. Otras ponen encima de la cabeza una columnita de un pie de altura y trenzan alrededor de ella el pelo y lo cubren con un velo negro.[7]

Figura 9. Cabeza de la Dama de Elche.

Figura 10. Parece que agrandar la cabeza mediante el peinado era una costumbre normal entre las poblaciones de finales de la Edad del Bronce de la vertiente atlántica de la Península Ibérica. Estela de Torrejón El Rubio II (según Almagro 1966).
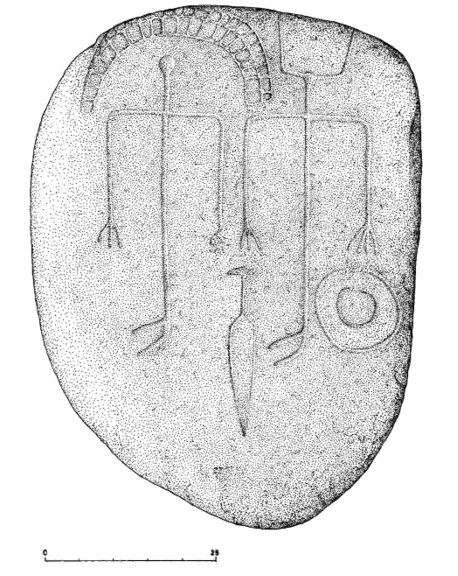
Figura 11. Estela de Almadén de la Plata (según García Sanjuán y otros 2006). La distinción de sexo entre el personaje masculino y el femenino queda atestiguada por las armas del primero, asociadas a la representación de la derecha, que se cubre con casco de cuernos y porta espada en su mano derecha y escudo en la izquierda. La compañera va tocada con una amplia diadema que le agranda la cabeza, un rasgo típico de neotenia.

Figura 12. Dama íbera sedente del Cerro de los Santos. De nuevo aparece aquí el característico engrosamiento de la cabeza como rasgo paidomórfico, esta vez en forma de tocado textil.
La evolución corporal se nutre de diversos motores que originan diversidad en la edad reproductora de los organismos. Como estamos viendo, uno de esos mecanismos fuente de variación en el adulto es la prolongación de las características somáticas y fisiológicas infantiles. Por eso, la ausencia en las mujeres de un cambio en el timbre de voz tan acusado como el que acontece en la pubertad de los varones puede de nuevo interpretarse como la permanencia de esos rasgos paidomórficos en la hembra, cosa grata por tanto al macho. Aunque hay que suponer tal característica en las mujeres protohistóricas hispanas, reconocerá con nosotros el lector que no contamos con ningún documento arqueológico probatorio de su existencia.
Una última peculiaridad en la que se acercan los patrones infantiles y los cánones de belleza atañe a los cuidados prestados por las personas mayores a los niños para el aseo corporal. La higiene propia y la del hogar, incluido el despiojamiento del otro, representan atenciones elementales facilitadas por muchos animales a su descendencia, norma que se incrementa en las especies cuya reproducción tiende a la estrategia k (pocos hijos y mucho empeño en sacarlos adelante). Algunas de estas acciones se relacionan, cuanto menos, con dos comportamientos sociales de los adultos: marcar la jerarquía dentro del grupo y normalizar el preámbulo de la unión sexual. Las aves suelen limpiar de excrementos sus nidos; por otra parte, se ha visto a algunas madres chimpancés asear a sus bebés con fibras vegetales humedecidas. También aquí algunos etólogos han observado una confluencia evolutiva, que tendría un vector en el acicalamiento de la infancia y otro en los cuidados corporales mutuos que se prestan los amantes (Eibl-Eibesfeldt 1995: 130-155). No es nuestra, por tanto, la idea de relacionar ambas conductas; pero sí la que propone hacer extensiva esta explicación a las tareas que buscan proporcionar a los adultos olores agradables mediante perfumes y ungüentos. Desde tal enfoque, no sería casual por tanto la concurrencia en el sarcófago femenino de Cádiz de dos características observadas en el cuidado que la mujer se prestaba a sí misma como táctica de engalanamiento: el peinado relativamente ostentoso que agranda la cabeza y el frasco de perfume que el personaje porta en su mano izquierda (fig. 13). Los muchos ungüentarios protohistóricos hispanos incitan a adentrase más por esta línea de trabajo, especialmente por la abundancia de esos botecillos en las sepulturas femeninas, pero retrasar esta moda a tiempos más antiguos requiere una previa investigación funcional de las vasijas aún no realizada. En cualquier caso, el mundo egipcio, en cuyas pinturas y relieves aparece representada con frecuencia la mujer con el denominado pseudocono de ungüento, puede servir como botón de muestra de esta clara relación entre el uso de perfumes y el erotismo (Padró 2007: 114).
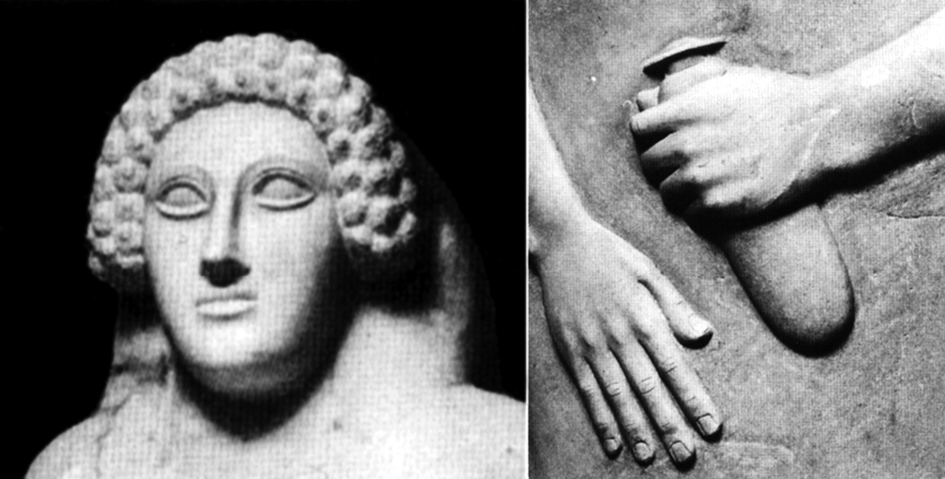
Figura 13. Sarcófago fenicio femenino de Cádiz. Presenta dos características típicas del exorno femenino: la amplitud del volumen de la cabeza logrado mediante el peinado y el uso de perfume, este último contenido en el alabastrón que lleva en la mano.
Por lo que hoy sabemos, la mujer antigua acaparó las normas de autoembellecimiento que concordaban con la emulación inconsciente de los rasgos paidomórficos. Una excepción hispana podría ser la institución cántabra de la covada. Desde el análisis evolucionista, esta costumbre dispondría de una explicación parecida a la que hemos trabajado en este artículo hasta ahora, explicación que soslaya, pero que no niega ni apoya, la clásica interpretación que la liga a una transición desde estructuras matriarcales al patriarcado (Caro Baroja 1976: 194; García y Bellido 1993: 207). Ya en época romana Estrabón (III, 4,17) mostró extrañeza ante el hecho de que el padre de la criatura permaneciera un tiempo en cama tras el parto, esencia de la costumbre. Tal asombro, que coincidiría también con el de cualquier occidental de hoy, emana de nuestra predisposición reciente a identificar formalmente alumbramiento y enfermedad, cosa que ha llevado a los países del primer mundo a parir casi siempre en centros hospitalarios. A causa del bipedismo, es vedad que para Homo sapiens este trance es más penoso que para otros mamíferos. Quizás por ello la selección natural ha primado las sensaciones dolorosas y la demanda de asistencia, porque ese proceder habría salvado de una muerte segura en muchos casos tanto a la madre como al bebé. La ayuda ajena ante tales señas de alarma repercute en el aumento numérico del grupo, y por tanto de sus genes y sus memes. Sin embargo, no ha sido siempre normal en la historia humana que se relacionen mentalmente parto y enfermedad (Rosenberg y Trevathan 2002: 62-67). Como consecuencia, parece poco probable que la permanencia en cama de los padres cántabros durante la temporada que seguía al nacimiento de su hijo pueda identificarse de forma directa con el hecho de que adquirieran desde ese momento la condición individual y social de enfermos. Nuestra interpretación propone más bien que se trata nuevamente de una emulación del comportamiento infantil, tal vez llevada a cabo ahora de forma consciente. Más que la situación de la madre, el padre remedaría el estado de desprotección del bebé. Se trataría, pues, de una solicitud gestual de atenciones que el marido lanzaba a su esposa, cuidados semejantes a los que la madre procuraba a la nueva criatura y con los que podrían verse reanudadas las relaciones sexuales del matrimonio.
Una reflexión darwiniana como la que aquí seguimos debería explicar, además de estos detalles, un asunto más general: por qué la convergencia evolutiva entre las características infantiles y las estrategias de obtención de pareja se centraron, con excepción de la covada, en la producción de semejanzas analógicas paidomórficas sólo en la mujer. Una respuesta posible diría que los papeles fisiológicos de género estuvieron implicados en la trama, pero faltan aún estudios etológicos sobre las culturas arcaicas que puedan satisfacer los pormenores de esta respuesta. Nuestra sospecha hipotética es que pudo ser relevante el ideal de compañero que de forma paralela desarrollaron desde tiempos ancestrales las mujeres. La búsqueda de un protector de la prole, sobre todo en ecosistemas inestables en los que se hacía necesaria la agresión para la defensa de los recursos, daría cuenta de la razón fundamental por la que las hembras encontraron especial atracción sexual en machos fuertes, curtidos y valerosos. La figura del guerrero adquiere de esta forma la categoría de arquetipo del compañero ideal. Se trataría por tanto de una evolución opuesta por completo a la femenina. Por eso al varón no le convenían los caracteres que van asociados a la infancia, siempre asimilables a situaciones de indefensión. En cualquier caso, no es posible pensar estas situaciones como estrategias masculinas conscientes y voluntarias, más propias de una visión lamarckiana de los cambios culturales. Una perspectiva darwinista sostendría más bien que las inclinaciones estéticas surgieron aquí, como cualquier mutación, también de manera espontánea, y que la posterior selección natural fue el mecanismo encargado de coaligar pautas culturales e intereses genéticos. En un proceso paralelo, la evolución dio paso en las correspondientes mujeres a una predilección por aquellos varones que mostraban esas características externas en mayor grado.
Si en las mujeres la relación analógica entre las estrategias sexuales y las características infantiles se ha perpetuado a lo largo de miles de años, convendríamos en que tal cooperación ha sido rentable para la reproducción de las poblaciones humanas que han contado con ella, o al menos más prolífica que otras conductas posibles. Pero la misma razón explicaría el fracaso de la covada y por tanto su final en la historia evolutiva de los grupos humanos de Hispania. Porque parece improbable, aun si se hubiesen producido plenas relaciones sexuales subsidiarias de las peticiones de atención por parte de los maridos encamados, que tales cópulas desembocaran en embarazos inmediatos al parto, sobre todo porque la calidad nutricional de las mujeres cántabras, normalmente baja en aquella época, habría impedido en ellas la ovulación y la lactancia de consuno. Por eso, si la covada no repercutió ni positiva ni negativamente en la cantidad de prole lograda por el grupo que la practicaba, quienes comenzaran tales usos habrían experimentado una mutación memética neutra, esto es, un cambio de costumbres sin consecuencias demográficas. Pero si esos posibles coitos provocaron infecciones o graves problemas de salud para las esposas al no estar separados lo bastante del parto, la covada acabó por convertirse en una mutación memética negativa. La selección natural debería haberla eliminado, como así hizo, por sus efectos deletéreos.
BELLEZA Y SALUD. OTRA ALIANZA EVOLUTIVA
Una máxima de los ganaderos españoles resume los lazos existentes entre una vigorosa salud y la belleza deseable en los rebaños domésticos: “dame gordura y te daré hermosura”. El refrán reúne estado físico y predilección estética, otra hipótesis bien trabajada por la biología evolutiva para dar explicación a las pautas de atracción entre sexos opuestos. Una salud robusta rara vez acompaña a un aspecto flaco o a una tez macilenta. Unas carnes rosadas y una mucosa labial colorida representan algunas de las garantías del buen estado de salud (Hamilton y otros 1990; Jablonski y Chaplin 2000). La apariencia y la estructura de la cara femenina suministran a menudo datos acerca del vigor del fenotipo completo, incluido el funcionamiento de las glándulas internas, pero también del genotipo. Estas cualidades se relacionan en especial con los marcadores hormonales, entre ellos con niveles altos en estrógenos y bajos en testosterona (Barber 1995; Johnston y Franklin 1993; Symons 1995; Thornhill y Gangestad 1996; Perret y otros 1994). De ahí que el semblante constituya todo un indicador de la situación física y, por ende, de la posición social[8]. Por esta correlación, la evolución ha seleccionado situaciones que hacen converger los intereses del genotipo con los gustos estéticos heredados por aprendizaje. Así, la búsqueda de pareja guiada por los cánones culturales de belleza transmitidos por la tradición ha constituido el marchamo primado por la evolución para que la relación sexual acabe en reproducción. En el varón, una constitución atlética, una buena estatura y un cuerpo musculoso garantizaban en la Iberia protohistórica el amparo de la institución familiar. En razón de tal presupuesto, la teoría evolutiva predice el éxito de los gustos estéticos oportunos, de forma que los memes más eficaces infundirían en las mujeres de la época la inclinación hacia varones con esa facha. En el sexo opuesto, el vocablo “gordura” que el dicho celebra quedará limitado a ciertos rasgos que acentúan las condiciones físicas necesarias para una feliz procreación: principalmente anchura de caderas, esteatopigia y tamaño de las mamas. Estos signos se plasman en las correspondientes «curvas», tan estimadas por los varones en la elección de compañera.
La cantidad y distribución de grasa en las caderas son sólo en parte resultado de la constitución ósea de la pelvis y del canal del parto, además de un buen indicador del funcionamiento hormonal (Domínguez-Rodrigo 2007: 31 y 33). Por tanto, expresan en cierto sentido lo fácil o complicado que será el alumbramiento cuando llegue. Los glúteos alojan depósitos de lípidos idóneos para etapas de hambruna y para atender al alimento extra que demanda el feto. Por eso el reparto corporal de grasa en las mujeres ha evolucionado como indicador de juventud, salud y fertilidad (Singh 1995). Finalmente, unas ubres como Dios manda certifican que la descendencia saldrá adelante. Por todas estas razones, los memes que, de manera velada o expresa, llevan al macho a elegir la compañera más apta posible para la reproducción, dispondrán de más posibilidades de supervivencia en la labor de criba de la selección natural. Lo mismo puede aseverarse si esas preferencias por determinado aspecto femenino están intervenidas por replicadores de efecto aplazado, es decir, que se activan cuando el varón adolescente agudiza su erotismo.
A propósito de las estelas grabadas de finales de la Edad del Bronce y del Hierro Antiguo, antes hemos analizado ya cómo los rasgos masculinos hacen con frecuencia alusión a cuerpos esbeltos y en cierta medida atléticos, por lo que no insistiremos más en el tema. Sí nos pararemos ahora, en cambio, en los principales caracteres femeninos, que cuentan con una nutrida documentación arqueológica. En alguna ocasión se ha argumentado que la ocultación de determinados miembros del cuerpo es directamente proporcional a su valor erótico. Este fenómeno se ha reconocido en muchas culturas, y permitiría defender que, en la Península Ibérica, hasta bien entrado el Holoceno las mamas podrían haber carecido de un elevado aprecio sexual. Aunque conocemos vestimentas de mujeres que cubren casi todo el cuerpo en momentos de la Edad del Cobre, son las representaciones protohistóricas de época prerromana las que detallan con cierta asiduidad mujeres vestidas desde el cuello para abajo (fig. 14). Aunque esto revela que los tabúes sexuales afectarían a gran parte del cuerpo femenino en las culturas que usaron esas imágenes, debemos reconocer nuestra precaria situación a la hora de indagar en este aspecto concreto, donde apenas si podemos ofrecer unas cuantas conjeturas. De hecho, desnuda nos ha llegado la imagen de bronce de la Astarté del Carambolo (fig. 15) y no puede deducirse de ello que los fenicios carecieran de ese tipo de prohibiciones[9]. Más oportuno parece asumir, tanto en este caso como en el de algunas figurillas calcolíticas anteriores que muestran cuerpos femeninos en cueros, que se trata de representaciones de divinidades, porque los dioses podían carecer del pudor típico de los humanos. Si la narración bíblica de la creación cuenta que Yahvé hizo al varón y a la varona a su imagen y semejanza, y que los puso en el paraíso desnudos (Génesis 1-2), esos textos orientales vendrían a apoyar de alguna forma esta característica de la divinidad. Tal vez por esta razón han surgido algunas dudas acerca de que las figurillas de mujeres obesas de algunas culturas neolíticas sean realmente representaciones de la Diosa Madre mediterránea, ya que pudieron haber contado en su origen con vestidos textiles sobrepuestos (Masvidal 2007: 212).

Figura 14. Dama del Cerro de los Santos. El vestido femenino cubría en época ibérica prácticamente todo el cuerpo.

Figura 15. Exvoto de bronce del Carambolo. Representa a Astarté entronizada.
Es evidente que los reclamos sexuales estudiados en este apartado no se parecen a las características infantiles. Por esta razón, no todas las estrategias pueden explicarse mediante la convergencia analógica con la peculiaridades neoténicas. En este caso se trata más bien de la coincidencia de memes y genes en una misma meta, su propia pervivencia. A fin de cuentas, ambos tipos de replicadores involucrados en la atracción intersexual ponen especial énfasis en las consecuencias reproductoras de la elección idónea de pareja, y fomentan de esta manera mecanismos de retroalimentación positiva que analizaremos a continuación con cierto detenimiento.
En algunos aspectos que pocos han leído desde un enfoque evolucionista, las imágenes antiguas de mujeres y de hombres muestran una imitación evidente de la realidad física. Un ejemplo concreto se refiere a la coloración de la piel. La femenina (más clara) y la masculina (más oscura) de los frescos egipcios o etruscos, pero también de la cerámica clásica griega, responden a un intento por parte de los correspondientes pintores de plasmar lo que sus ojos percibían en la vida real, hecho también observado discretamente en algunos ambientes hispanos de época prerromana (fig. 16). De hecho, los matices cromáticos de la epidermis humana en relación con la dualidad de sexos, expresados ya en las pinturas neolíticas saharianas mucho antes que en las faraónicas, cretenses o pompeyanas de época posterior (Camps 1974: 245), tienen que ver con adaptaciones milenarias promovidas por la selección natural, sobre todo para la optimización de la relación beneficios/perjuicios de la luz solar, con importantes repercusiones en la diversidad racial y en el dimorfismo sexual (Jablonski y Chaplin 2000). Concretamente en las mujeres, este hecho se vincula estrechamente con la mayor o menor capacidad para sintetizar vitaminas y compuestos químicos implicados en una correcta gestación. Así, las pieles más blancas incrementan el ácido fólico, un compuesto que los ginecólogos de hoy proporcionan de inmediato a las mujeres nada más comenzar el embarazo para evitar males en el feto. No se trata por tanto de una moda artística la costumbre de los pintores de entonces de representar a cada sexo con un color distintivo. Ni debería caber extrañeza alguna en el hecho de que, en la franja subtropical mediterránea, donde la luz solar es más débil que en el segmento ecuatorial terrestre, se prodiguen memes que hacen apetecible para el hombre la mujer de piel clara. Es de esta forma como se explica desde el darwinismo la nívea hermosura de las princesas homéricas, caso por ejemplo de Nausícaa (Odisea, canto VI), la tez resplandeciente de la dama egipcia descrita en el Papiro de Chester Beatty (Padró 2007: 113) y, por supuesto, los piropos dedicados en las inscripciones funerarias romanas a las muchachas que morían antes de lo esperado: blancas como el mármol[10].

Figura 16. Aunque son muy pocos los documentos que pueden usarse para el estudio del color de la piel en la Hispania prerromana, algunas representaciones muestran al hombre (derecha) más oscuro que a la mujer (izquierda). La representación femenina pertenece a un vaso cerámico de La Alcudia, la masculina a uno de Ampurias.
LA PERVIVENCIA COMO PREMIO
Aún habiendo dedicado la mayor parte de este trabajo a la sexualidad en general y, más específicamente, a la selección sexual, desde un enfoque evolucionista resulta inexcusable tratar el tema del éxito reproductivo por dos razones fundamentales: por las consecuencias que sobre la demografía tienen las distintas prácticas sexuales y por las repercusiones que sobre las pautas culturales ejercen las consecuencias reproductoras de la sexualidad. En cualquier caso, seremos especialmente breves en este apartado por no alargar excesivamente el trabajo; en definitiva, es justamente el triunfo reproductivo el tema que ha abarcado mayor espacio en los estudios evolucionistas sobre sexualidad.
En cualquier sistema dado hay una constante de mecanismos que operan en dos sentidos básicos. Hay primero una creación persistente de desorden y desestabilización, entendidos estos factores como el surgimiento de novedosas circunstancias que ensanchan el campo de la diversidad. Existen después otros procesos de estabilización puntual y conservación. En esta otra tarea, el motor principal es la selección natural, que consigue dar una determinada dirección a todos y cada uno de los elementos que conforman un sistema mediante mecanismos de elección y preservación. En el fondo, no estamos expresando de esta forma más que las dos partes de la segunda ley de la termodinámica, porque la primera tendencia es gratis y espontánea y la segunda está necesitada de añadir energía externa al sistema, justo lo que afirma esta regla física (Atkins 1992). Pues bien, ambos mecanismos –la variabilidad y la selección natural– ocupan un lugar central en el desarrollo de cualquier ecosistema de nuestro planeta. En primer lugar, la diversidad viene dada por la misma reproducción. Evidentemente, hay sistemas que permiten una mayor variabilidad que otros. La reproducción asexual en general produce descendientes con códigos génicos idénticos a los que portan los predecesores. En este caso apenas se da variación. En cambio, la reproducción sexual permite una amplia gama de diversidad[11]. En segundo lugar, la selección natural se encarga mediante mecanismos de criba, de descarte o preservación en definitiva, de encauzar las directrices de individuos y grupos.
Como ya hemos avanzado, después de leer An essay on the principle of population, obra que T.R. Malthus había publicado en 1798, Darwin comprendió que el mecanismo por el que se generalizan en la naturaleza determinadas transformaciones surgidas por azar tiene que ver directamente con las diferencias reproductoras de individuos y poblaciones (Ruiz y Ayala 1999: 306-309). Así, toda transformación tendente a acrecentar la estirpe se incorporará a las futuras generaciones en proporción progresiva. De esta forma, se da cabida a la evolución sin necesidad de que se produzcan modificaciones de los ecosistemas o del ambiente. Por eso la evolución no puede ser definida, según se ha entendido casi siempre, como simple adaptación, pese a que, al asumir tal mecanismo, el propio Darwin ajustó su idea de cómo la diversidad puede operar a favor de que las especies se acomoden al medio. A pesar de lo cual, tuvo que ahondar en la concepción monista del individuo, es decir, en la aceptación de que cuerpo y conducta se manifiestan siempre como un todo, la unidad sobre la que opera la selección.
La conclusión principal que desde el enfoque evolucionista podemos aplicar a los grupos humanos de la Protohistoria ibérica es que su comportamiento más común y persistente es tan producto de la selección natural como su carga genética y su fenotipo. Habría que añadir, incluso, que cualquier cambio conductual que incrementara la demografía tenía amplias posibilidades de convertirse en moda entre la gente que vio surgir dicha mutación memética. Por tanto, la generalización al conjunto de la población de las estrategias de cortejo que antes hemos analizado puede tener que ver directamente con esas posibilidades de contribuir al auge demográfico. Desde un enfoque darwinista, el proceso se inicia siempre en individuos concretos, que por adoptar las nuevas pautas ven aumentado su éxito en la captación de pareja, y subsidiariamente su prole. A través de la herencia cultural, y gracias a la tendencia innata a imitar, los descendientes adquieren de sus progenitores la innovación. A la larga, dicha mutación memética positiva acaba por caracterizar a todo el linaje, para más tarde pasar a convertirse en norma de toda una población mayor, sea tribu, pueblo o cultura. De esta forma, y a modo de cuello de botella (Ayala 1994: 103-104) –eso sí, sin necesidad de reducción drástica de la población–, la competencia memética, iniciada en principio entre muy pocos individuos, ha ensanchado su campo de actuación al cabo de generaciones, de forma que comienza pronto a enfrentar conjuntos formados por más de una persona, primero bajo la modalidad de la «selección de parentela» y más tarde en la denominada «selección de grupo». El mecanismo funciona como si la selección natural operase eligiendo entre elementos conformados internamente por conjuntos de individuos caracterizados por comportamientos homogéneos. Es así como comprendemos desde una interpretación darwiniana la sustitución paulatina de unas poblaciones por otras y por tanto de unas culturas por otras, cada una con sus correspondientes esquemas de pensamiento y con sus pautas de conducta propias. Este es el ejemplo aducido normalmente para explicar, en tiempos muy anteriores a los que ahora analizamos, la sustitución de los neandertales por los humanos modernos[12].
La fórmula que permite comprender científicamente este crecimiento diferencial y, más tarde o más temprano, el reemplazo de unas poblaciones y/o culturas por otras, se basa en los modelos de reproducción que los biólogos han denominado estrategias k y r. La reproducción bajo la selección de k origina poca descendencia e invierte paralelamente mucho esfuerzo en su crianza. La reproducción bajo la selección de r produce en cambio camadas numerosas, pero los progenitores apenas si dedican energía a sacarlas adelante. Ambas tácticas suponen valores relativos, nunca categorías absolutas. De esta forma, dentro incluso de una misma especie, unas poblaciones pueden inclinarse a k y otras a r. Es la elección de una ruta u otra la causa de importantes consecuencias en la demografía genética, pero también en la memética. De ello se deduce que, a la hora de caracterizar culturalmente a un grupo social concreto o a una etnia determinada, se verán incrementados numéricamente aquellos patrones etológicos que contribuyan a colocar más descendencia en edad reproductora para perpetuar la cadena. La estrategia r es sin duda un modelo reproductivo que tiende a aumentar la población, de forma que debe ser tenida en cuenta a la hora de analizar los cambios en la secuencia cultural de la Prehistoria Reciente y Protohistoria hispanas (fig. 17).
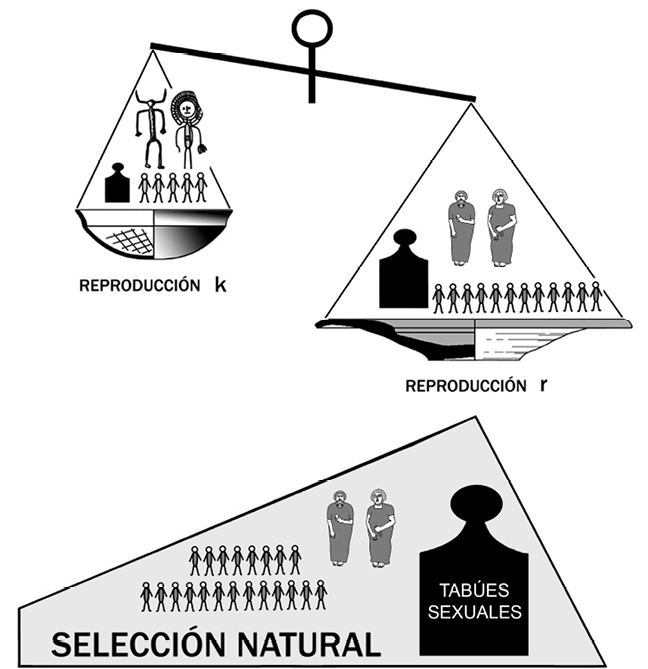
Figura 17. Posiblemente, la abundancia de fenicios en la Hispania protohistórica señalada por Estrabón no se deba tanto a una llegada masiva de población cananea cuanto a su alto índice de crecimiento demográfico. A este éxito reproductivo pudieron contribuir diversos factores, entre los que no pueden olvidarse los preceptos legales y morales sobre la sexualidad, así como la costumbre de casarse las mujeres a edad muy temprana.
La relación de la reproducción diferencial de individuos y poblaciones con la sexualidad es por tanto un factor de capital importancia a la hora de analizar la evolución cultural de las comunidades humanas. Según estos principios teóricos del darwinismo, toda idea o comportamiento (todo meme o complejo de memes interrelacionados) que contribuya a aumentar la demografía, seguramente será incorporado al acerbo cultural de una población por simple intervención de la selección natural. Es de esta forma como la propuesta evolucionista predice que cualquier tabú sexual dispondrá de un campo abonado para arraigar y propagarse si la prohibición no afecta de forma negativa a las consecuencias reproductoras de la sexualidad, pero sobre todo si las aumenta[13]. En la tradición judeocristiana de Occidente, tal fenómeno está reflejado a la perfección en el sexto mandamiento de la ley mosaica, que demoniza las relaciones sexuales «impuras», siendo así que las «puras» o lícitas vienen representadas por las practicadas dentro de la pareja legal y destinadas a procrear (o que no impidan el embarazo). He aquí concentradas dos necesidades básicas para la perpetuación generacional: eliminar interferencias en la producción de prole con el rechazo expreso de la sexualidad voluntariamente yerma, y dotar a las crías de un medio seguro (matrimonio-familia) que garantice su llegada hasta la edad fértil. Dada la antigüedad de este precepto veterotestamentario, cabe sospechar la existencia posible de normas parecidas en otras culturas de entonces. En las hispanas, esta conducta pudo estar presente al menos entre las poblaciones de origen fenicio. De hecho, aunque no hemos podido constatar prohibiciones sexuales expresas en los textos religiosos cananeos, como ocurre por el contrario en el mundo hebreo con la abominación yahvista del onanismo (Génesis 38) y en Egipto con el recelo hacia la homosexualidad (Padró 2007: 120), los documentos orientales del segundo milenio a.C. hablan de relaciones amorosas entre las propias divinidades que, ofrecidas a los humanos como guía arquetípica de lo que deben hacer, se limitan la mayor parte de las veces al coito y acaban siempre en fecundación de la diosa primero y, finalmente, en parto[14]. Así, por ejemplo, el ciclo menor de Baal, en el que éste copula como dios-toro con una novilla-diosa que engendrará el consiguiente ternero:
“[Baal (?)] apretó y agarró su vulva,
[Anat (?)] apretó y agarró sus testículos.
[Y Ba]al desfalleció (?) por el becerro (?),
[por el morlaco de] la Virgen Anat.
[Al abrazarse/besarse (?) hubo] concepción y parto
[con la ayuda (?)] de la banda de las Kotharot:
[un hijo nació (?) a] la Virgen Anat,
[progenie a] Baal el Todopoderoso.”
(KTU 1.11)[15]
Hoy podemos aceptar que todo este mundo religioso ugarítico de finales del segundo milenio a.C., caracterizado por unos fuertes lazos entre sexo y reproducción (Cunchillos 2007), fue prácticamente el mismo que, comenzado el milenio siguiente, los fenicios llevaron hasta la Península Ibérica. Los estudiosos de la religión feniciopúnica reconocen de hecho que los credos cananeos de la Edad del Hierro son una herencia palmaria de los del Bronce Tardío siropalestino (Marín 2002: 16). Numerosos documentos arqueológicos han venido a certificar, por lo demás, los vínculos directos entre esos dioses semitas y los bóvidos ya en las colonias fenicias hispanas. Es de esta forma, en calidad de arquetipo del coito divino primordial, como puede interpretarse la escena de cópula de los relieves de la torre funeraria de Pozo Moro (fig. 18), transcrita siempre en la literatura arqueológica como hierogamia propiciatoria de la fecundidad (López Pardo 2006: 96-100).

Figura 18. Hierogamia de Pozo Moro. El apareamiento divino sirve a los humanos como ejemplo del correcto proceder sexual.
Pero lo que la arqueología y la antropología física están comenzando a conocer de la Protohistoria ibérica con cierta profundidad son algunos aspectos relativos a la demografía a partir del registro funerario, tema del que queremos ofrecer una pincelada para acabar. Por eso, en lo tocante a la reproducción y a la sexualidad, son elocuentes, entre otros muchos datos, los publicados por M.E. Aubet sobre los túmulos del Hierro Antiguo de Setefilla (Aubet 1995: 402). En tales enterramientos, el análisis de las incineraciones ha mostrado una esperanza de vida para la población femenina en torno a los veinte años. Se ha achacado esta corta cifra a posibles complicaciones de sobreparto, lo que estaría demostrando la edad en que las mujeres accedían a las relaciones sexuales. En este sentido, hay que tener presente la imposibilidad que tenía entonces la cultura de alargar la vida fértil de las mujeres; pero también la facultad que poseía por contra para hacer coincidir el matrimonio con el inicio de ésta, con lo que se aprovechaban las máximas posibilidades fisiológicas para la procreación. Se diría que, ante la necesidad de descendencia por parte de unas familias que tenían como principal mecanismo de producción el trabajo manual, los memes implicados en el establecimiento de lazos matrimoniales habrían experimentado una evolución conducente a promover lo que los biólogos han denominado «estrategia pesimista», aquella que, ante una corta esperanza de vida, que en Setefilla rara vez superaba los treinta años y sólo en el caso de los varones, inducía a las mujeres a un pronto casamiento. Un perfil casi idéntico al que presenta este yacimiento español constituía la realidad del mundo egipcio de época faraónica (Padró 2007: 107). Por eso mismo, sexualidad y reproducción estaban estrechamente relacionadas también en la cultura griega, tanto como en otras muchas sociedades primitivas (Plácido 2007: 188).
UNA ALIANZA COPULATIVA
En relación con el tema trabajado en el presente artículo, puede llegarse a la conclusión de que genes y memes cooperaron durante miles de años para proporcionar a la selección natural la materia prima necesaria que definió la vida sexual de las culturas arcaicas mediterráneas. Por tanto, para la gente que habitó la Iberia protohistórica, las costumbres relativas al sexo y sus consecuencias sobre la reproducción conforman una herencia biológica marcada por pautas heredadas difícilmente eludibles[16].
En el ideal de belleza que arraigó en aquellas gentes del mundo antiguo, semejante a los cánones que en gran medida aún perfilan nuestros gustos actuales, convergieron intereses genéticos y meméticos, somáticos y conductuales. No es necesaria una explicación que enfrente a genes y memes por ver quién controla ya la evolución, porque dicha conclusión parte de visiones no monistas –y por tanto acientíficas– del ser humano. La colaboración entre ambos tipos de replicadores aparece en cambio con profusión. Es más, tampoco sería necesario optar por una de las dos líneas que hemos trabajado para comprender por qué sendero evolutivo caminaron las predilecciones estéticas de la gente protohistórica. Así, es posible que la hipótesis que ve en estas normas el plagio inconsciente de indicadores somáticos de buena salud haya acertado en muchas de sus propuestas, pero no es menos cierto que, en cualquier caso, esas mismas características pueden considerarse convergencias paidomórficas. En consecuencia, más que una simple disyuntiva entre las dos, una explicación evolucionista más compleja y completa, darwinista en cualquier caso, podría perfectamente asumir una relación copulativa entre ambas: «y» mejor que «o».
BIBLIOGRAFÍA
ACOSTA, P. y PELLICER, M. (1990): La Cueva de la Dehesilla (Jerez de la Frontera). Las primeras civilizaciones productoras en Andalucía occidental. CSIC - Centro de Estudios Históricos Jerezanos, Jerez de la Frontera.
ALEXANDER, R. (1994): Darwinismo y asuntos humanos. Salvat., Barcelona.
ALMAGRO, M. (1966): Las estelas decoradas del suroeste peninsular (Bibliotheca Praehistorica Hispana VIII). CSIC, Madrid.
ALVAR, J.; MUÑIZ, E. y DE LA VEGA, T. (2007): “Restricciones sexuales en los cultos mistéricos” en S. Celestino (ed.), La imagen del sexo en la Antigüedad: 225-250. Tusquets, Barcelona.
ANDERSSON, M. (1994): Sexual selection, Princeton University Press.
ARSUAGA, J.L. y MARTÍNEZ, I. (1998): La especie elegida. La larga marcha de la evolución humana. Temas de Hoy, Madrid.
ATKINS, P.W. (1992): La segunda ley. Prensa Científica, Barcelona.
AUBET, M.E. (1995): “Aproximación a la estructura social y demográfica tartésica”, Tartessos 25 Años Después. 1968-1993, Jerez de la Frontera: 401-409. Ayuntamiento de Jerez de la Frontera, Jerez de la Frontera.
AYALA, F.J. (1994): La teoría de la evolución. De Darwin a los últimos avances de la genética. Temas de Hoy, Madrid.
BARBER, N. (1995): “The evolutionary psychology of physical attractiveness: sexual selection and human morphology”, Ethology and Sociobiology 16: 395-424.
BERMEJO, J.C. (1984): “«La calvicie de la luna»: Diodoro Sículo XXXIII, 7, 5 y la posible existencia de un nuevo mito turdetano”, Gerión 2: 181-191.
BERMEJO, J.C. (1986): Mitología y mitos de la Hispania prerromana, vol. 2. Akal, Madrid.
BLACKMORE, S. (2000): La máquina de los memes. Paidós, Barcelona.
BLANCO, A. y CORZO, R. (1981): “Der neue anthropoide Sarkophag von Cádiz”, Madrider Mitteilungen 22: 236-243.
CAMPS, G. (1974): Les civilisations préhistoriques de l’Afrique du nord et du Sahara. Doin, Paris.
CANT, J. (1991): “Hypothesis for the evolution of the human breasts and buttocks”, American Naturalist 117: 199-204.
CARO BAROJA, J. (1946): Los pueblos de España. Ensayo de etnología. Barna, Barcelona.
CARO BAROJA, J. (1976): Los pueblos de España, vol. I. Istmo, Madrid.
CELESTINO, S. (2001): Estelas de guerrero y estelas diademadas. La precolonización y formación del mundo tartésico. Bellaterra, Barcelona.
CUNCHILLOS, J.L. (2007): “La sexualidad en la cultura semítica noroccidental: Ugarit”, en S. Celestino (ed.), La imagen del sexo en la Antigüedad: 125-144. Tusquets, Barcelona.
DARWIN, C. (1871): The descent of man, and the selection in relation to sex. John Murray, London.
DAWKINS, R. (1979): El gen egoísta. Las bases biológicas de nuestra conducta. Salvat, Barcelona.
DAWKINS, R. (2007): El espejismo de Dios. Espasa, Barcelona.
DE HOZ, J. (1991): “Las lenguas célticas peninsulares”, en Los Celtas en la Península Ibérica. Revista de Arqueología (nº especial): 36-41.
DEL OLMO, G. (1998): Mitos, leyendas y rituales de los semitas occidentales. Trotta – Universidad de Barcelona, Madrid.
DOMÍNGUEZ-RODRIGO, M. (1994): El origen del comportamiento humano. Tipo, Madrid.
DOMÍNGUEZ-RODRIGO, M. (2004): El origen de la atracción sexual humana. Akal, Madrid.
DOMÍNGUEZ-RODRIGO, M. (2007): “El sexo en tiempos de Adán y Eva. Una visión evolucionista de la sexualidad humana”, en S. Celestino (ed.), La imagen del sexo en la Antigüedad: 25-48. Tusquets, Barcelona.
EIBL-EIBESFELDT, I. (1995): Amor y odio. Historia natural del comportamiento humano. Salvat, Barcelona.
ELDREDGE, N. y GOULD, S.J. (1972): “Punctuated equilibria: an alternative to phyletic gradualism”, en T.J.M. Schopf (ed.), Models in Paleobiology: 82-115. Freeman and Cooper, San Francisco.
ELIADE, M. (1972): El mito del eterno retorno, Alianza, Madrid.
ENDLER, J. y BASOLO, A.L. (1998): “Sensory ecology, receiver biases and sexual selection”, Trends in Ecology and Evolution 13 (19): 415-420.
ESCACENA, J.L. (2000): “Applications of evolutive archeology: migrations from Africa to Iberia in the Recent Prehistory”, en A. Arnáiz-Villena (ed.), Prehistoric Iberia. Genetics, anthropology and linguistics: 125-162. Kluwer Academic-Plenum Publishers, New York.
ESCACENA, J.L. (2002): “Darwin (in)comprendido. Evolución humana y Prehistoria en la tradición académica andaluza. Breve análisis de una paradoja”, en M. Belén y J. Beltrán (ed.), Arqueología fin de siglo. La arqueología española de la segunda mitad del siglo XIX (I Reunión Andaluza de Historiografía Arqueológica) (Spal Monografías III): 205-225. Universidad de Sevilla, Sevilla.
ESCACENA, J.L. (2005): “Darwin y Tartessos”, en S. Celestino y J. Jiménez (ed.), El Periodo Orientalizante (Anejos de Archivo Español de Arqueología XXXV): 189-219. CSIC, Mérida.
FERNÁNDEZ MARTÍNEZ, C. (2002): “CLE 1996 (Epitafio De Julia Paula): comentario filológico”, Habis 33: 301-323.
FERRER, E. y DE LA BANDERA, M.L. (2007): “Santuarios, aldeas y granjas: el poblamiento durante el Bronce final y el Período orientalizante”, en E. Ferrer (ed.), Arqueología en Marchena. El poblamiento antiguo y medieval en el Valle medio del río Corbones: 39-79. Universidad de Sevilla, Sevilla.
GARCÍA RIVERO, D. (2004): “Prehistoria y evolución: reflexiones sobre la secuencia ecológico-cultural holocénica en el mediodía ibérico”, Spal 13: 9-34. http://dx.doi.org/10.12795/spal.2004.i13.01
GARCÍA SANJUÁN, L.; WHEATLEY, D.W.; FÁBREGA, P.; HERNÁNDEZ, M.J. y POLVORINOS, A. (2006): “Las estelas de guerrero de Almadén de la Plata (Sevilla). Morfología, tecnología y contexto”, Trabajos de Prehistoria 63 (2): 135-152.
GARCÍA Y BELLIDO, A. (1967): Veinticinco estampas de la España antigua, Espasa Calpe, Madrid.
GARCÍA Y BELLIDO, A. (1993): España y los españoles hace dos mil años según la Geografía de Estrabón, Espasa Calpe, Madrid.
GONZÁLEZ MORALES, M.R. (2007): “La imagen del sexo en el Paleolítico”, en S. Celestino (ed.), La imagen del sexo en la Antigüedad: 49-66. Tusquets, Barcelona.
GOULD, S.J. (1994): El pulgar del panda. Reflexiones sobre historia natural y evolución. Crítica, Barcelona.
GOULD, S.J. (2004): La estructura de la teoría de la evolución. Tusquets, Barcelona.
Hamilton, W.D.; Axelrod, R. y Tanese, R. (1990): “Sexual reproduction as an adaptation to resist parasites (A rewiew)”, Proceedings of the National Academy of Science (USA) 87:.3566-3573.
HERNANDO, A. (2007): “Sexo, género y poder. Breve reflexión sobre algunos conceptos manejados en la Arqueología del Género”, Complutum 18: 167-174.
IZQUIERDO, R. y LÓPEZ JURADO, S. (1998): “Estela de guerrero de El Coronil (Sevilla), Spal 7: 177-182. http://dx.doi.org/10.12795/spal.1998.i7.11
JABLONSKI, N.G. y CHAPLIN, G. (2000): “The evolution of human skin coloration”, Journal of Human Evolution 39 (1): 57-106.
JOHNSTON, V. S. y FRANKLIN, M. (1993): “Is the beauty in the eye of the beholder?”, Ethology and Sociobiology 14: 183-199.
JONES, D. (1996): “An evolutionary perspective on physical attractiveness”, Evolutionary Anthropology 5: 97-109.
LÓPEZ PARDO, F. (2006): La torre de las almas. Un recorrido por los mitos y creencias del mundo fenicio y orientalizante a través del monumento de Pozo Moro (Anejos de Gerión X). Universidad Complutense, Madrid.
MARÍN, M.C. (2002): “En torno a las fuentes para el estudio de la religión fenicia en la Península Ibérica”, en E. Ferrer (ed.), Ex Oriente Lux: Las religiones orientales antiguas en la Península Ibérica: 11-32. Universidad de Sevilla-Fundación El Monte, Sevilla.
MARTÍ, B. y HERNÁNDEZ, M.S. (1988): El neolític valencià. Art rupestre i cultura material. Diputación de Valencia, Valencia.
MARTÍNEZ, G. y LANATA, J.L. (ed.) (2002): Perspectivas integradoras entre arqueología y evolución: teoría, métodos y casos de aplicación. Olavarría, Incuapa.
MASCHNER, H.D.G. (ed.) (1996): Darwinian archaeologies. Plenum, New York.
MASVIDAL, C. (2007): “Bases para una nueva interpretación sobre las mujeres en la Prehistoria”, Complutum 18: 209-215.
MORRIS, D. (1995): El mono desnudo. Plaza y Janés, Barcelona.
MUSCIO, H.J. (2006): “Una aproximación evolutiva a la complejidad y al orden social durante el período temprano a través del estudio de representaciones rupestres. El caso de la Quebrada de Matancillas, Puna Argentina”, Estudios Atacameños 31: 1-28.
NICOLINI, G. (1977): Bronces ibéricos. Gustavo Gili, Barcelona.
O’BRIEN, M.J. y LYMAN, L. (2000): Applying evolutionary archaeology. A systemcatic approach. Plenum, New York.
PADRÓ, J. (2007): “La sexualidad en Egipto”, en S. Celestino (ed.), La imagen del sexo en la Antigüedad: 107-124. Tusquets, Barcelona.
PERICOT, L. (1979): Cerámica ibérica. Polígrafa, Barcelona.
Perrett, D.I.; May, K.A. y Yoshikawa, S. (1994): “Facial shape and judgments of female attractiveness”, Nature 368: 239-242.
PLÁCIDO, D. (2007): “El sexo en la sociedad griega: la paideía, los rituales y los mitos”, en S. Celestino (ed.), La imagen del sexo en la Antigüedad: 187-205. Tusquets, Barcelona.
PRESEDO, F. (1973): La Dama de Baza. Ministerio de Cultura, Madrid.
PURVES, D.; BEAU LOTTO, R. y NUNDY, S. (2003): “Por qué vemos lo que vemos”, Investigación y Ciencia 323: 56-64.
QUEROL, M.A. (2001): Adán y Darwin. Síntesis, Madrid.
RINDOS, D. (1990): Los orígenes de la agricultura. Una perspectiva evolucionista. Bellaterra, Barcelona.
ROSENBERG, K.R. y TREVATHAN, W.R. (2002): “La evolución del parto humano”, Investigación y Ciencia 304: 62-67.
RUEDA, C. (2007): “La mujer sacralizada: La presencia de las mujeres en los santuarios (lectura desde los exvotos de bronce ibéricos)”, Complutum 18: 227-235.
RUIZ, R. y AYALA, F. (1999): “El núcleo duro del darwinismo”, en F. Glick y otros (ed.), El Darwinismo en España e Iberoamérica: 299-323. Universidad Nacional Autónoma de México - CSIC, Madrid.
RUIZ-GÁLVEZ, M. (1992): “La novia vendida: orfebrería, herencia y agricultura en la Protohistoria de la Península Ibérica”, Spal 1: 219-251. http://dx.doi.org/10.12795/spal.1992.i1.11
RUIZ-GÁLVEZ, M. (1995): “A modo de epílogo. La Ría de Huelva: conclusiones y perspectivas”, por M. Belén y otros, en M. Ruiz-Gálvez (ed.), Ritos de paso y puntos de paso. La Ría de Huelva en el mundo del Bronce Final europeo (Extra 5 de Complutum). 157-166. Universidad Complutense, Madrid.
RUSE, M. (2001): El misterio de los misterios ¿Es la evolución una construcción social? Tusquets, Barcelona.
SCHULTEN, A. (1952): Fontes Hispaniae Antiquae. Universidad de Barcelona, Barcelona.
SHENNAN, S. (2002): Genes, memes and human history. Darwinian archaeology and cultural evolution. Thames & Hudson, London.
SINGH, D. (1995): “Female health, attractiveness, and desirabilit fo relationships: Role of breast asimmetry and waist-to-hip ratio”, Ethology and Sociobiology 16: 465-481.
SYMONS, D. (1995): “Beauty in the adaptation of the beholder: the evolutionary psychology of human female sexual attractiveness”, en P.R. Abramson y S.D. Pinker (ed.), Sexual Nature / Sexual Culture: 80-118. University of Chicago Press, Chicago.
TOBEÑA, A. (2005): Mártires mortíferos. Biología del altruismo letal. Un itinerario por el cerebro de los suicidas atacantes. Universidad de Valencia, Valencia.
THORNHILL, R. y GANGESTAD, S.W. (1996): “The evolution of human sexuality”, Trends in Ecology and Evolution 11: 98-102.
WILSON, E.O. (1980): Sociobiología. La nueva síntesis. Omega, Barcelona.
WRIGHT, S. (1945): “Tempo and mode in evolution. A critical review”, Ecology 26: 415-419.
Recepción: 19 de diciembre de 2007. Aceptación: 16 de septiembre de 2008
[1] Esto se ha afirmado especialmente para el marxismo en relación con las sociedades de cazadores-recolectores (cf. Domínguez-Rodrigo 2004: 5). Nosotros añadiríamos más: si el materialismo dialéctico no explica una parte del todo, tampoco explica el todo. Cualquier teoría científica, o sirve para el análisis global, o queda infravalorada epistémicamente frente a otras que sí cumplen este requisito (Ruse 2001: 48).
[2] Parafraseando lo afirmado por S.J. Gould (2004: 444) sobre Hugo de Vries, se diría que muchos evolucionistas actuales son antidarwinistas a pesar suyo.
[3] En el siguiente apartado entraremos de lleno en la evolución cultural como expresión de la conducta aprendida, cosa que se trasmite por emulación memética. Los memes serían los replicadores básicos del comportamiento no heredado por vía genética y de la ideología que lo sustenta. Parece toda una sarcástica paradoja que R. Dawkins, el creador de la voz meme con la que aquí nos sentimos tan cómodos, sea el autor de una reciente obra eumemésica cuyo valor científico es verdaderamente lamentable (cf. Dawkins 2007).
[4] «Ruido de fondo» debe entenderse aquí como plesiomorfias, es decir, caracteres primitivos, aquellos que compartimos con otros muchos animales por herencia común. En el terreno de la sexualidad, un ejemplo podría ser la inexistencia en la hembra humana de periodo de celo estacional. Puede que tal rasgo, semejante por ejemplo al de los bonobos, sea el resultado de nuestro origen ancestral en zonas de selva tropical. Frente a otros medios, en estos ecosistemas muy húmedos escasean los episodios cíclicos de falta de alimento vegetal (Domínguez-Rodrigo 1994: 27-28). Tal hecho hace innecesaria la sincronización del destete con un momento determinado del año en que impere la abundancia. Aunque en algunas especies actuales de primates la ausencia de celo estacional pueda ser una homología, es decir, una semejanza originada ya en un ascendiente común, también compartimos dicha peculiaridad con aves y otros muchos seres vivos que medran en esos ambientes selváticos, lo que debe interpretarse en este último caso como una analogía adquirida por convergencia. Esta interpretación sobre la ausencia de un periodo concreto de celo en la mujer no anula otras interpretaciones que ven en tal característica la condición necesaria para que el varón permanezca siempre junto a ella, posible origen para muchos de la familia humana (Domínguez-Rodrigo 2007: 26)..
[5] Una traducción completa del texto en García y Bellido (1967: 79-80).
[6] Si la selección actúa en la misma trayectoria, organismos distintos suelen desembocar en situaciones de apariencia similar. Los ecosistemas helados han dado lugar de esta forma a especies que han tendido a ser rechonchas y a estar bien dotadas de grasa externa, dos remedios contra las frías temperaturas. Esto se puede observar en los osos, en las focas y en los pingüinos, sin que tal solución suponga parentesco cercano entre esos animales. En teoría evolutiva, dichos rasgos convergentes se denominan analogías. Sin embargo, los parecidos menos superficiales, es decir, aquellos que se manifiestan en la estructura ósea por ejemplo, y que incumben al diseño interno y global de dos o más organismos, constituyen homologías. Las semejanzas se deben en este caso a una herencia compartida. Los dos tipos de afinidades han sido trabajados en biología humana tanto en su versión somática (cf. Arsuaga y Martínez 1998: 144-145) como conductual (p.e. Eibl-Eibesfeldt 1995: 40-47).
[7] Traducción de A. Schulten (1952: 114).
[8] Es evidente que la posición social se exhibe también con la cantidad y calidad de adornos que se llevan en la cabeza. Por eso, la Dama de Elche puede ser definida en términos populares como “un buen partido”. Precisamente el atesoramiento de joyas y de lujosas vajillas en tiempos protohistóricos ha sido puesto en relación con el establecimiento de lazos matrimoniales, bien con la dote bien con el precio de la novia (cf. Ruiz-Gálvez 1992).
[9] La Astarté del Carambolo no solo está desnuda. Su figura muestra senos simétricos y firmes, y en general el cuerpo de una mujer joven. La predilección masculina por pechos grandes y estables se ha generalizado, así parece, por su valor para diferenciar a las mujeres más jóvenes de las más ancianas, sobre todo por la fragilidad de las estructuras de suspensión de las mamas tras repetidas secuencias de preñez y lactancia (Cant 1991). Elegir a una compañera de corta edad era y es mayor garantía de éxito reproductivo si no concurren otras variables contrarias a la procreación, sean éstas de tipo físico o cultural.
[10] Valga como ejemplo el epitafio de Julia Paula, equiparada al mármol de Paros (Fernández Martínez 2002: 304-305).
[11] Aunque la replicación genética sexual es digital, es decir, con un grado de perfección bastante preciso, normalmente la prole suele habilitar algún tipo de cambio respecto a sus progenitores, básicamente debido a la recombinación cromosómica. Varios estudios han achacado el éxito de las comúnmente denominadas “especies superiores” al tipo de reproducción que ostentan, la sexual. Ésta posibilita además la entrada en escena de unos mecanismos que fortalecen tales efectos, los que engloba la selección sexual. En definitiva, este sistema reproductivo permite generalmente una mayor rapidez en el ritmo de cambio de las especies y una mayor difusión en número de individuos.
[12] Para explicar en parte tal sustitución se he echado mano del énfasis puesto por Homo sapiens en las connotaciones sexuales y reproductoras de la figura femenina superopaleolítica (González Morales 2007: 61-63).
[13] Para esta hipótesis representa un reto la explicación de las limitaciones sexuales que imponían ciertas religiones antiguas de origen oriental extendidas por el mundo romano, sobre todo porque muchas de ellas, como la voluntaria castración de los galli de Cibeles, implicaban la imposibilidad de originar estirpe propia (Alvar y otros 2007). Aunque la ausencia de datos de este tipo para la Protohistoria ibérica nos libera de ofrecer aquí un tratamiento amplio del tema, convendría no olvidar que la selección de grupo ha actuado históricamente promoviendo el beneficio genético y/o memético de la comunidad mediante la renuncia a la reproducción individual. Hoy, el caso más extremo de este fenómeno es la autoinmolación que llevan a cabo los terroristas suicidas (Tobeña 2005).
[14] Para la mentalidad del hombre arcaico, las narraciones míticas cumplen normalmente la función de establecer la norma correcta de comportamiento según fue al principio de los tiempos (in illo tempore), hasta el extremo de que el concepto de pecado va asociado a la realización de actos que carecen de referencia arquetípica en el plano divino o en el proceder primordial de los antepasados divinizados o héroes (Eliade 1972: 13-52).
[15] Edición y traducción de G. del Olmo (1998: 132-133).
[16] Entendemos el término “biológico” como la unidad de lo somático-fisiológico y lo conductual. Desde nuestra posición teórica, también la cultura es biología. Es más, la Arqueología Evolutiva sostiene que la tecnología se transforma, como los seres vivos, por selección natural, en tanto que acrecienta o disminuye en los organismos que la usan la adaptación –y por tanto la fitness– a los ecosistemas en los que medran.