DOI: https://dx.doi.org/10.12795/rea.2024.i48.01
Formato de cita / Citation: Poma-Sarango, M.A., Poma-Sarango, D., Guaman, L., Chuncho, C.G., Juela, O., & Benavides-Silva, C. (2024). Potential distribution of ecosystems in the Southern Zone of Ecuador: modeling from a correlative approach. Revista de Estudios Andaluces,(48), 8-32. https://dx.doi.org/10.12795/rea.2024.i48.01
Correspondencia autores: cesar.benavidez@unl.edu.ec (César Benavides-Silva)
Marco Antonio Poma-Sarango
marco.poma@unl.edu.ec  0009-0001-6149-2403
0009-0001-6149-2403
Diego Poma-Sarango
diego.poma@unl.edu.ec 0000-0002-8622-6216
Lizbeth Guaman
lizbeth.guaman@unl.edu.ec 0009-0000-5063-8545
Carlos Guillermo Chuncho
carlos.chuncho@unl.edu.ec 0009-0001-9299-0595
Oscar Juela
oscar.juela@unl.edu.ec 0000-0002-3560-8206
César Benavidez-Silva
cesar.benavidez@unl.edu.ec 0000-0002-9822-4103
Centro de Investigaciones Territoriales. Universidad Nacional de Loja, Ecuador.
Ciudad Universitaria Guillermo Falconí, Loja. Ecuador.
|
INFO ARTÍCULO |
RESUMEN |
|
|
Recibido: 17-10-2023 Revisado: 29-12-2023 Aceptado: 26-01-2024 PALABRAS CLAVE Cambio climático Modelización Ecosistemas Biomod2 Zona Sur del Ecuador |
El cambio climático es uno de los factores que están afectando a los ecosistemas terrestres a través de la variabilidad climática global y, al actuar de manera sinérgica con otros factores estresantes, se potencian sus efectos sobre los ecosistemas pudiendo afectar directamente a la diversidad biológica. En consecuencia, hemos analizado la distribución potencial de los ecosistemas de la Zona Sur del Ecuador bajo un escenario de cambio climático futuro (RCP 6.0 2080), de importancia ecológica, social y económica para las poblaciones aledañas a los ecosistemas. Para ello se utilizó los modelos de distribución potencial (SDM) implementados en Biomod2 4.0, que utiliza un conjunto de métodos y herramientas relevantes para el tratamiento de una serie de incertidumbres metodológicas relacionadas con el modelado de distribuciones, así como la capacidad de construir rápidamente modelos individuales o combinarlos de diferentes maneras mediante ensambles, relacionando variables biofísicas (topográficas y climáticas) con los puntos de presencia/ausencia de los ecosistemas de la Zona Sur. Los resultados indican que la temperatura sufriría un incremento para todos los ecosistemas, mientras que la precipitación en algunos ecosistemas de pisos altitudinales altos, como Páramo y Bosque Montano Alto decrecería, diferente a lo que ocurriría con otros ecosistemas en pisos altitudinales más bajos, donde la precipitación mostraría un incremento, mientras que en los pisos altitudinales más altos, de manera general, la precipitación aumentaría. Esta variabilidad climática posiblemente genere nuevas configuraciones climáticas y los ecosistemas en respuesta tiendan a reducirse o expandirse, cambiando su rango de distribución. |
|
|
KEYWORDS |
ABSTRACT |
|
|
Climate Change Modeling Ecosystems Biomod2 Southern region of Ecuador |
Climate change is one of the factors affecting terrestrial ecosystems through global climate variability, and by acting synergistically with other stressors, its effects on ecosystems are enhanced, potentially directly affecting biological diversity. Consequently, we have analyzed the potential distribution of ecosystems in the Southern Zone of Ecuador under a future climate change scenario (RCP 6.0 2080), which is ecologically, socially, and economically important for populations surrounding the ecosystems. To do this, we used potential distribution models (SDM) implemented in Biomod2 4.0, which uses a set of relevant methods and tools for dealing with a series of methodological uncertainties related to distribution modeling, as well as the ability to quickly build individual models or combine them in different ways through ensembles, relating biophysical variables (topographic and climatic) with the presence/absence points of ecosystems in the Southern Zone. The results indicate that the temperature would increase for all ecosystems, while precipitation in some high-altitude ecosystems, such as Paramo and Montane Forest, would decrease, unlike what would happen with other ecosystems at lower altitudes, where precipitation would show an increase (and, in the rest of the ecosystems, in general, it would show an increase). This climate variability may generate new climate configurations and ecosystems may tend to shrink or expand in response, changing their distribution range. |
El clima es el resultado de un proceso de complejas interacciones entre factores atmosféricos, geográficos y meteorológicos, que inciden sobre los procesos ecológicos, económico y socio-productivos a nivel del global (Samaniego-Rojas et al., 2015). A lo largo de la historia del planeta, el clima ha estado en constante variación (IPCC, 2014), obedeciendo a ciclos dinámicos naturales en un principio (Benavidez-Silva et al., 2021), sin embargo en el último siglo el ser humano ha tomado protagonismo y es considerado como responsable de manera parcial o total por su contribución en el incremento de gases de efecto invernadero (GEI) (IPCC, 2014).
El cambio climático de origen antrópico es cada vez más vigente (Malhi et al., 2020), y afecta el equilibrio energético del sistema climático del planeta (IPCC, 2008), lo que ha ocasionado un incremento de la temperatura media de la superficie terrestre en ~1 °C en relación con la temperatura media de los años 1850-1900 (Dagtekin et al., 2020). Por ello, es considerado una amenaza global, cuyos efectos pueden ser cada vez más severos (Weiskopf et al., 2020), afectando a la salud física-mental de las personas (Parry et al., 2019); así como, poner en riesgo la biodiversidad actual del planeta (Correa-Lima et al., 2019). Las anomalías en los patrones de precipitación y temperatura a nivel global (Busch et al., 2012), principales efectos evidenciados a raíz del cambio climático (Colwell et al., 2008; Hylander et al., 2022), impactan negativamente a los ecosistemas terrestres a escala global (Dagtekin et al., 2020), obligando a las especies a modificar sus límites geográficos e impulsar la redistribución de la vida sobre el planeta (Ashraf et al., 2017; Pecl et al., 2017). Algunos organismos podrían evolucionar (ecológica o genéticamente) para adaptarse a estos cambios (Malhi et al., 2020; Thomas, 2020), mientras que el resto podría desaparecer (extinción) (Dagtekin et al., 2020).
Los ecosistemas naturales representan la principal fuente proveedora de recursos genéticos y servicios ecosistémicos, que permiten el desarrollo de la humanidad (Aguirre et al., 2017), sin embargo, el cambio global y el incremento de las presiones antrópicas, ocasionan transiciones críticas en la estructura y función de los ecosistemas (Rocha, 2022). El cambio climático podría alterar las condiciones ambientales de los distintos ecosistemas en respuesta a las actividades antrópicas (Benavidez-Silva et al., 2021), acelerando la pérdida de especies animales y vegetales, y con ello el deterioro de los ecosistemas, siendo más vulnerables aquellos que han sido alterados por las actividades humanas (CEPAL, 2017; Quesada-Quirós et al., 2016). El cambio en los regímenes de precipitación y temperatura, provocaría un desplazamiento de los ecosistemas hacia altitudes mayores, debido al incremento de la evapotranspiración (Colwell et al., 2008), lo que supone mayores impactos para los ecosistemas y especies asociadas, distribuidos en rangos altitudinales más elevados (Aguirre et al., 2015).
En la Zona Sur del Ecuador, conformada por las provincia de Loja, Zamora Chinchipe y El Oro, los cambios de uso de suelo, producto del crecimiento urbano, la deforestación, la minería y la expansión de la frontera agrícola, históricamente son la causa de la fragmentación de los ecosistemas, siendo los bosques secos los ecosistemas más amenazados debido a los procesos de conversión agrícola que han sufrido en los últimos años (Aguirre et al., 2015; Manchego et al., 2017; Tapia-Armijos et al., 2015). Los estresores climáticos y antrópicos han influenciado de manera negativa sobre los ecosistemas de la Zona Sur del Ecuador, convirtiéndola en una región vulnerable a los efectos del cambio climático (Aguirre et al., 2015).
En este contexto, la interacción de los patrones climáticos con otros factores de estrés, como los cambios en los usos del suelo, podría tener un impacto significativo en el equilibrio natural de los ecosistemas terrestres de las provincias de Loja, Zamora Chinchipe y El Oro en el futuro. Con el objetivo de comprender los patrones espaciales y temporales que rigen la distribución de los seres vivos en el planeta, se utilizaron modelos correlativos, específicamente Modelos de Distribución de Especies (SDM). Estos modelos fueron empleados para prever posibles zonas de distribución de los ecosistemas (Pliscoff & Fuentes-Castillo, 2011), proporcionando así información valiosa sobre la potencial respuesta de estos ecosistemas al cambio climático (Ashraf et al., 2017; Guisan & Zimmermann, 2000), las actividades humanas (Benavidez-Silva et al., 2021) y así, poder determinar las variaciones espaciales y temporales que sufrirían los ecosistemas en diferentes escenarios climáticos para prever su correcta gestión (Gaglio et al., 2019).
La comprensión de cómo el clima ha evolucionado en décadas recientes es crucial para anticipar los futuros cambios en la estructura de la biodiversidad (Carroll et al., 2017). En este estudio, se modelaron los ecosistemas terrestres de las provincias de Loja, Zamora y El Oro para el período 1970-2080, utilizando enfoques correlativos y considerando el escenario climático IPCC-AR5-RCP 6.0. La aplicación de los SDM en Biomod2 4.0 (Thuiller et al., 2022) permitió relacionar variables biofísicas con la ocurrencia/ausencia de los ecosistemas (Benavidez-Silva et al., 2021; Brun et al., 2020; Hao et al., 2019). En concreto, el estudio se propone evaluar las zonas de distribución potencial de los ecosistemas en la Zona Sur hacia el año 2080 bajo un escenario de cambio climático, específicamente para identificar las variables determinantes y aplicar modelos correlativos para prever la distribución futura de estos ecosistemas.
Esta investigación fue desarrollada en el espacio geográfico que abarca toda la Zona Sur del Ecuador (3° 02’ 52’’ S y 03° 02’ 41’’ S, y entre 78° 21’ 58’’ W y 78° 21’ 35’’ W) (figura 1), comprendida por las provincias de Loja, Zamora Chinchipe y El Oro. Esta región se distribuye en ~27.500 km2, equivalentes a ~11% del territorio nacional (Aguirre et al., 2015). El Ecuador debido a su ubicación geográfica presenta diversos tipos de climas y microclimas que generan zonas con altos niveles de biodiversidad (Samaniego-Rojas et al., 2015), como la Zona Sur del Ecuador, la cual presenta una gran diversidad climática, multiétnica y biológica, esta última se ve potenciada por la influencia de algunos factores como su ubicación geográfica, presencia de cuencas hidrográficas, diversidad de pisos altitudinales, formación fitogeográfica Huancabamba, presencia de valles secos interandinos y la presencia de grupos étnicos (Aguirre et al., 2015), la que se caracteriza por su gran diversidad de especies y endemismo (Brehm et al., 2008), está situada dentro de hotspots de biodiversidad (Andes Tropicales y Tumbes-Choco-Magdalena) (Tapia-Armijos et al., 2015), incluyendo valles secos, páramos, bosques nublados y bosques amazónicos (Brehm et al., 2008, Garzón-Santomaro et al., 2019) alcanzando altitudes de hasta 3.880 m s.n.m (Cueva, 2012).
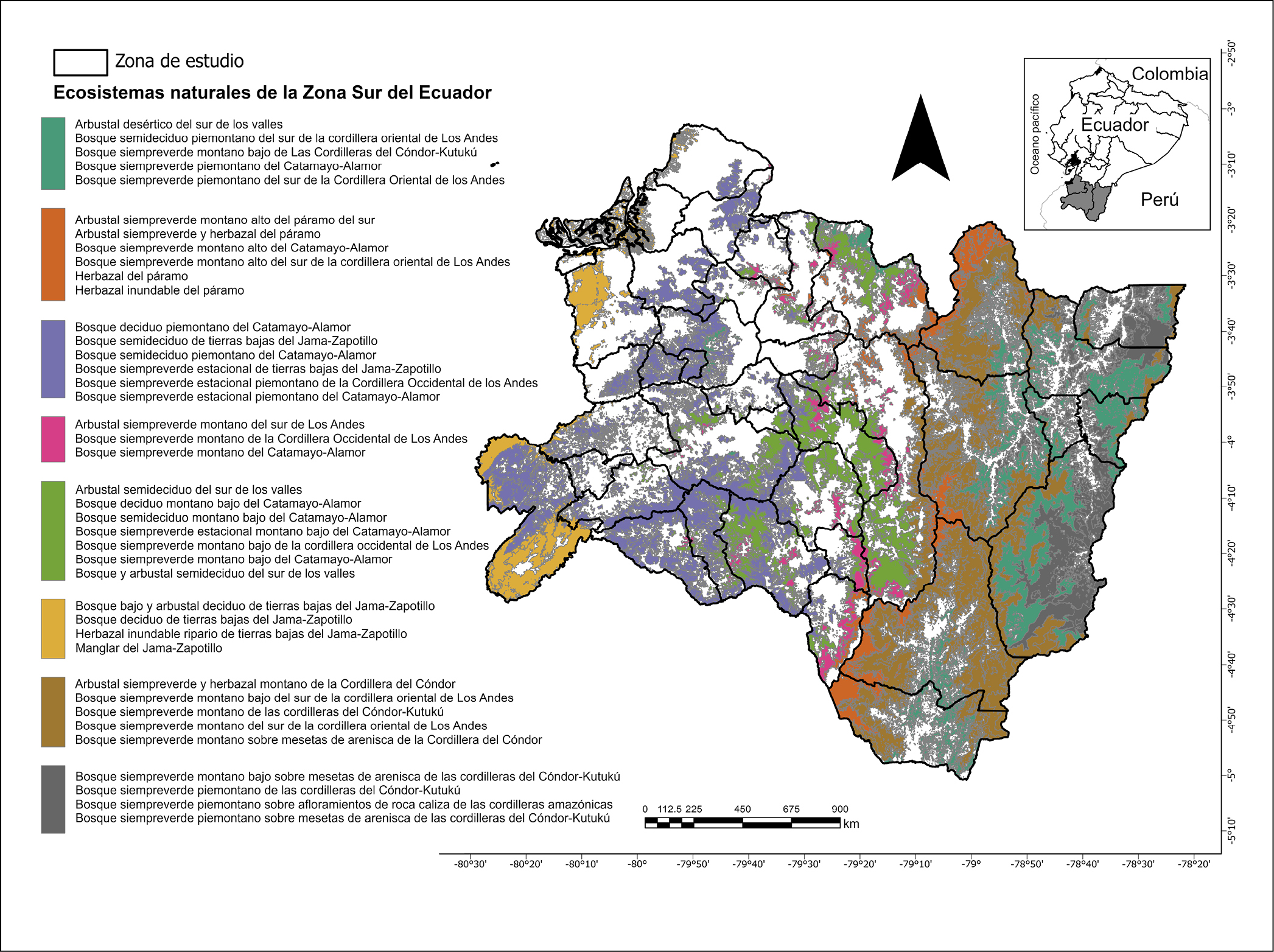
Figura 1. Zona Sur del Ecuador: mapa resumen de la distribución de los remanentes de los ecosistemas naturales provinciales. Fuente: (MAE, 2013). Elaboración propia.
Los ecosistemas naturales presentes en esta región se ven afectados por las variaciones interanuales del clima, que a su vez son determinados por factores geográficos y meteorológicos (latitud, orografía) y la presencia del océano pacífico (Samaniego-Rojas et al., 2015). La flora es una de las más ricas y diversas del mundo, la cual varía en función de los diferentes climas de la región (Lozano, 2002), que de manera general son: a) clima megatérmico seco a semihúmedo, las temperaturas promedio son superiores a los 22 °C y los niveles de precipitación oscilan entre 500 a 1.000 mm; b) clima ecuatorial mesotérmico semihúmedo a húmedo, se caracteriza por inviernos relativamente secos debido a la pérdida de humedad que sufren las masas de aire al chocar con los flancos de la cordillera. Las precipitaciones anuales promedio varían desde los 600 a 2.000 mm, con valores que llegan hasta los 6.700 mm anuales en las partes más altas de la cordillera, las temperaturas fluctúan entre los 12 – 22 °C ; c) clima tropical megatérmico muy húmedo: clima muy característico de la Amazonía ecuatoriana (Samaniego-Rojas et al., 2015).
La información ecológica requerida para el proceso de modelización corresponde a los datos de presencia de los ecosistemas modelados. Las presencias fueron obtenidas para las 40 formaciones vegetales o ecosistemas presentes en las provincias de Loja, Zamora Chinchipe y El Oro, de acuerdo a la clasificación del Ministerio del Ambiente Agua y Transición Ecológica (MAATE) (MAE, 2013), los mismos que se encuentran distribuidos en diferentes pisos bioclimáticos y con una fisiografía diferente a la del resto del país (Vazquez et al., 2005). Los puntos de ocurrencia de los ecosistemas (presencia) así como las ausencias son ambientalmente representativos y precisos para la escala de trabajo elegida (Pliscoff & Fuentes-Castillo, 2011).
Los datos de presencia fueron generadas considerando un mínimo de puntos entre 30 y 50, necesarios por cada ecosistema, para obtener modelos estadísticamente válidos (Kadmon et al., 2003) y para evitar la generación de modelos mal evaluados (Pliscoff & Fuentes-Castillo, 2011; Ren et al., 2016). Para obtener las presencias de los ecosistemas se siguió el siguiente criterio: para ecosistemas cuya distribución en el perfil altitudinal comprenda un rango entre 0 y 1.000 metros, el número de presencias generado fue de 50; entre 0 y 2.000 metros, el número de presencias generado fue de 100; y, entre 0 y ≥ 3.880 metros, el número de presencias generado fue de 150. Como los ecosistemas son unidades mutuamente excluyentes y sustitutos geográficos, la presencia de un determinado ecosistema se registró como (1) mientras que la ausencia se registró como (0) (Anjos & de Toledo, 2018) o lugares donde presumiblemente no se encuentran distribuidos los ecosistemas. Al no contar con datos de ausencias reales, se generaron puntos aleatorios (pseudoausencias), en la misma proporción para todos los ecosistemas (Benavidez-Silva et al., 2021; De Pando & De Giles, 2007), sin considerar el área de distribución observada de cada ecosistema; se generaron 10.000 puntos. Los datos de presencia y pseudoausencia de los ecosistemas fueron seleccionados aleatoriamente y georreferenciados en coordenadas geográficas con ayuda del software ArcGIS Pro 2.8.4.
Las variables ambientales implementadas en la presente investigación corresponden a variables bioclimática y variables topográficas. En esta investigación empleamos las 19 variables climáticas obtenidas del portal web Climatologies at high resolution for the earth’s land surface areas (CHELSA) (https://chelsa-climate.org/), a una resolución de 30 segundos (~1km en el Ecuador). Los periodos de tiempo que se propuso evaluar están basados en un escenario actual que representan un periodo de tiempo de 50 años (1970 – 2020) y un escenario climático futuro que corresponden a las extrapoladas con los modelos CIMP5 y MIROC5 para el 2080 respectivamente (promedio 2061 – 2080) bajo los modelos de concentración de gases de efecto invernadero RCP6.0 (Gang et al., 2017). Así mismo, se emplearon variables topográficas como hillshade, slope y aspect, derivadas del modelo digital de elevaciones (DEM), provisto por Worldclim (https://www.worldclim.org/data/index.html), basado en el SRTM (Farr et al., 2007) a una resolución espacial de 1 km, las cuales según Pliscoff & Fuentes-Castillo (2011), son de gran utilidad para obtener modelos de mayor ajuste y detalle.
Los modelos de distribución potencial de ecosistemas se generan usando dos tipos de fuentes de información: datos de presencia/ausencia de ecosistemas a modelar, así como las variables biofísicas para la Región Sur del Ecuador (Benavidez-Silva et al., 2021; Pliscoff & Fuentes-Castillo, 2011). El proceso de modelización de la distribución potencial se realizó en Bomod2 v 4.0, implementado en el software R (Thuiller et al., 2022), que se basa en el ensambles de diferentes técnicas de modelización (Pliscoff & Fuentes-Castillo, 2011), minimizando así las incertidumbres en la predicción de los modelos (Anjos & de Toledo, 2018) y que son usados para explicar la distribución de las especies o ecosistemas en distintas escalas espaciales y temporales (Fletcher & Fortin, 2018). En algunas investigaciones se ha demostrado un gran desempeño de la modelización correlativa de especies o ecosistemas en Biomod2 (Anjos & de Toledo, 2018; Benavidez-Silva et al., 2021; Ren et al., 2016), se pueden combinar para modelar la biodiversidad siguiendo la estrategia de predecir primero, ensamblar después (Ren et al., 2016). La modelización se ejecutó utilizando 10 métodos diferentes como: Generalized Linear Models (GLM); Generalized Additive Models (GAM); Generalized Boosted Models (GBM); Random Forests (RF); Multivariate Adaptative Regression Spline (MARS); Maximum Entropy Models (Maxent); Artificial Neural Networks (ANN); Surface Range Envelope (SER); Classification tree analysis (CTA); Flexible Discriminant Analysis (FDA) (Anjos & de Toledo, 2018; Hao et al., 2019; Pliscoff & Fuentes-Castillo, 2011). La modelización consta de 3 etapas importantes como calibración, evaluación y modelación/proyección (figura 2) (Benavidez-Silva et al., 2021).

Figura 2. Esquema metodológico para la modelización de ecosistemas. Fuente: elaboración propia.
Una vez obtenidas las observaciones para cada ecosistema, se determinaron las medidas correlativas entre las variables bioclimáticas y topográficas para eliminar la multicolinealidad (Benavidez-Silva et al., 2021), evitando que los modelos se sobreajusten (Ren et al., 2016) y minimizar la incertidumbre de la predicción del modelo (Anjos & de Toledo, 2018). Los predictores altamente correlacionados fueron eliminados del modelo usando la correlación por pares de Pearson con un umbral de 0,8 (Benavidez-Silva et al., 2021; Brun et al., 2020; Ren et al., 2016) y el método de Variance Inflation Factor (VIF) donde se consideró que los valores superiores a 5 en primera instancia o 10 en un segundo análisis que podrían indicar multicolinealidad entre variables, lo que causaría errores de predicción (Atauchi et al., 2020; Benavidez-Silva et al., 2021). Para evaluar la calidad de los predictores que mejor describen la presencia de los ecosistemas en el espacio climático, se calibró los modelos mediante el procedimiento de validación cruzada, que consiste en dividir los datos de presencia de manera aleatoria en dos subconjuntos, el 70% para calibrar y el 30% restante para evaluar las predicciones del modelo (Benavidez-Silva et al., 2021; Thuiller, 2003).
Es fundamental evaluar la precisión predictiva (o desempeño) de los modelos de distribución (Allouche et al., 2006), para ello, es fundamental elegir métricas adecuadas que permitan caracterizar adecuadamente la confiabilidad de los resultados generados, en este estudio empleamos dos métodos estadísticos para evaluar el rendimiento de los modelos: True Skill Statistics (TSS) y Receiver Operating Characteristic (ROC (AUC)) (Anexo 1), que se basan en la sensibilidad y especificidad (Anjos & de Toledo, 2018; Benavidez-Silva et al., 2021; Pliscoff & Fuentes-Castillo, 2011). Tanto ROC y TSS utilizan estas métricas para evaluar los SDM correlativos. Los índices se representan por medio de una gráfica que mide el equilibrio entre la sensibilidad (proporción de presencias verdaderas respeto al número total de presencias proyectadas) y especificidad (proporción de presencias falsas respecto al número total de ausencias proyectadas) en función del umbral de clasificación (Harrison et al., 2007).
En esta etapa se calculó la contribución de cada predictor bioclimático en cada modelo que permite explicar los patrones de la variables de respuesta que varía entre cero (menor importancia) y 1 (mayor importancia) (Anjos & de Toledo, 2018). Para esta investigación se consideró un umbral de 0,8 como métrica para evaluar los modelos, los modelos que superaron esté umbral se consideraron para los ensambles finales (Anjos & de Toledo, 2018; Benavidez-Silva et al., 2021).
La proyección en el espacio geográfico es el paso final en el proceso de modelización (Pliscoff & Fuentes-Castillo, 2011). Cada modelo se ejecutó para cada ecosistema, se comparó y se usó para derivar predicciones espaciales sobre los datos originales. La última etapa de la modelización de ecosistemas considera los modelos con el mejor poder predictivo para la proyección en el espacio geográfico y climático (Pliscoff & Fuentes-Castillo, 2011), la misma que será implementada luego de haber calibrado y evaluado el desempeño de los modelos y se implementará un umbral específico que permita identificar los valores inferiores al umbral como ausencia de ecosistemas y valores superiores al umbral serán considerados como presencias (Benavidez-Silva et al., 2021).
Luego de modelar el ecosistema actual, se implementó un método de ensamble, incluyendo todos los modelos de Biomod2, permitiendo reducir la incertidumbre bajo el poder predictivo (<0,8 en el TSS y ROC). Los modelos con el mejor poder predictivo se consideraron para el ensamble de los modelos, considerando los umbrales individuales de cada técnica, los cuales nos podrían permitir obtener un resultado más robusto, estos modelos individuales fueron proyectados posteriormente al espacio geográfico y climático actual y luego se proyectaron al espacio geográfico y climático futuro para el año 2080, implementado una Trayectoria de Concentración Representativa (RCP, por sus siglas en inglés), el RCP 6.0, propuesta por el IPCC en el Quinto Informe de Evaluación del Cambio Climático (AR5) (IPCC, 2014),considerando las mismas variables predictivas establecidas en la etapa de calibración (Benavidez-Silva et al., 2021).
Para el año 2080 de los ecosistemas de la Zona Sur del Ecuador (Anexo 2), bajo la influencia del cambio climático, considerando el escenario de emisión RCP6.0, se logró identificar las distintas variables que se emplearon para el proceso de modelización individual de cada ecosistema, las cuales se pueden evidenciar en Anexo 3, donde se identificó que las variables ambientales son relativamente divergentes de un ecosistema a otro, aun si comparten pisos bioclimáticos similares. Bajo este contexto y de acuerdo a los resultados obtenidos, se determinó que las variables topográficas son las utilizadas con mayor frecuencia en el proceso de modelización, mientras que, de las variables climáticas, las que tuvieron mayor frecuencia de uso fueron la Bio2, Bio7, Bio18, Bio4 y Bio12 (Anexo 3).
Con las proyecciones realizadas a futuro de los 40 ecosistemas de la Zona Sur del Ecuador, se procedió a realizar un mapa de áreas de recambio (figura 3), en donde se puede identificar zonas en donde los ecosistemas potencialmente podrían compartir el espacio geográfico, es decir, que los ecosistemas podrían sobreponer su distribución en distintas zonas con similitud climática y geográfica en el futuro, provocando un comportamiento difuso y una limitante metodológica de los modelos correlativos de distribución potencial. Las zonas de color rojo nos indica cuales serían las áreas de mayor recambio o sobreposición de condiciones climáticas y topográficas que podrían darse en el futuro, indica que podría existir el potencial para el establecimiento de hasta 12 ecosistemas; mientras que el color azul, indica las áreas donde posiblemente no se dé la sobreposición de condiciones climáticas y; por lo tanto, solo permita el establecimiento de ecosistemas con una limitada idoneidad climática.

Figura 3. Áreas de recambio de la Zona Sur del Ecuador bajo la influencia del cambio climático (escenario RCP 6.0). Fuente: resultados del proceso de modelización. Elaboración propia.
Las zonas de menor convergencia corresponden a los límites provinciales, las costas de la provincia de El Oro, el cantón Loja dentro de la provincia de Loja y gran parte de la provincia de Zamora Chinchipe. En la provincia de El Oro en las costas del cantón Santa Rosa, Huaquillas y Machala, presentarían las condiciones para que se desarrollen ecosistemas del piso bioclimático Tierras bajas, entre ellos el Manglar del Jama-Zapotillo (BsTc05), Bosque deciduo de tierras bajas del Jama-Zapotillo (BdTc01), Bosque bajo y arbustal deciduo de tierras bajas del Jama-Zapotillo (BdTc02) y Bosque siempreverde estacional de tierras bajas del Jama-Zapotillo (BeTc02). Dentro de la provincia de Zamora Chinchipe y parte de la zona centro y Sur del cantón Loja, se desarrollarían condiciones que permitirían el establecimiento desde 1 hasta 7 u 8 ecosistemas aproximadamente, entre ellos ecosistemas pertenecientes a los pisos bioclimáticos Montano, Piemontano y Montano bajo, como Bosque siempreverde montano de la cordillera occidental de los Andes (BsMn03), Bosque y arbustal semideciduo del sur de los valles (BmBn02), Bosque siempreverde piemontano sobre mesetas de arenisca de las cordilleras del Cóndor-Kutukú (BsPa04), Bosque siempreverde piemontano sobre afloramientos de roca caliza de las cordilleras amazónicas (BsPa03), Bosque siempreverde piemontano de las cordilleras del Cóndor-Kutukú (BsPa02), Bosque siempreverde montano de las cordilleras del Cóndor-Kutukú (BsMa02), Bosque siempreverde montano bajo las cordilleras del Cóndor-Kutukú (BsBa02), Bosque siempreverde y herbazal montano de la cordillera del cóndor (AsMa01) y Arbustal siempreverde montano del sur de los andes (AsMn02). Algunos ecosistemas presentarían presencias relativamente pequeñas distribuidos dentro del área de estudio, tales como Bosque siempreverde montano bajo sobre mesetas de arenisca de las cordilleras del Cóndor-Kutukú (BsBa03), Bosque siempreverde montano bajo de la cordillera occidental de los andes (BsBn04), Bosque siempreverde estacional piemontano de la cordillera occidental de los andes (BePn01), Bosque siempreverde montano alto del Catamayo-Alamor (BsAn04), BeTc02 en la provincia de El Oro y Arbustal siempreverde y herbazal del páramo (AsSn01) en la provincia de El Oro y Loja (Zaruma, Chila y Saraguro respectivamente).
Las zonas de mayor convergencia dentro de la Zona Sur están comprendidas por parte del territorio de la provincia de Loja, El Oro y el oeste del cantón Yacuambi de Zamora Chinchipe. Se estima que parte de los cantones Espíndola, Quilanga, Loja, Catamayo, Saraguro, Macara, Sozoranga, Portovelo, Zaruma, Chila y Atahualpa, presentarían condiciones tanto climáticas como topográficas aptas para el establecimiento de entre 5 hasta 12 ecosistemas. En la zona Norte, en los cantones Saraguro, Chila, Atahualpa, Zaruma, Portovelo y Yacuambi, los ecosistemas que podrían converger son Bosque deciduo montano bajo del Catamayo-Alamor (BdBn01), Arbustal desértico del sur de los valles (AdBn01), AsSn01, Bosque siempreverde estacional montano bajo del Catamayo-Alamor (BeBn01), Arbustal siempreverde montano alto del páramo del sur (AsAn01), Bosque siempreverde montano alto del sur de la cordillera oriental de los andes (BsAn02), Bosque semideciduo piemontano del Catamayo-Alamor (BmPn01), Bosque siempreverde estacional piemontano del Catamayo-Alamor (BePn02), Bosque siempreverde montano bajo del sur de la cordillera oriental de los andes (BsBn02), Bosque siempreverde piemontano del sur de la cordillera oriental de los andes (BsPn04), Bosque siempreverde estacional piemontano de la cordillera occidental de los andes (BePn01), BsBn04 y BsBa03, pertenecientes a los pisos bioclimáticos Montano alto, Montano alto superior, Montano bajo y Piemontano. En la zona centro y Sur de la Zona Sur, en los cantones Catamayo, Gonzanamá, Loja, Espíndola, Quilanga y Loja principalmente y una pequeña parte de los límites con Chinchipe y Palanda, los ecosistemas que podrían coincidir son Bosque semideciduo piemontano del sur de la cordillera oriental de los andes (BmPn02), Bosque semideciduo montano bajo del Catamayo-Alamor (BmBn01), Bosque siempreverde montano del Catamayo-Alamor (BsMn04), Bosque siempreverde montano del sur de la cordillera oriental de los andes (BsMn02), Herbazal del páramo (HsSn02), Bosque deciduo piemontano del Catamayo-Alamor (BdPn01), Bosque siempreverde montano bajo del Catamayo-Alamor (BsBn05), Arbustal siempreverde y herbazal montano de la cordillera del cóndor (AsMa01), BmBn02 y BsMn03, pertenecientes a los pisos bioclimáticos Piemontano, Montano bajo, Montano y Montano alto superior.
De los 40 ecosistemas analizados, 2 serían gravemente afectados por el cambio climático, provocando su desaparición dentro de la Zona Sur del Ecuador. Estos ecosistemas son Herbazal inundable del páramo (HsSn04) y Bosque siempreverde montano sobre mesetas de arenisca de la cordillera del cóndor (BsMa01), estos ecosistemas potencialmente podrían migrar latitudinalmente hacia el norte principalmente (~500 a 1.000 km) (ocupando zonas del norte de Ecuador y territorio colombiano), y en menor proporción hacia el sur (Perú).
Los cambios en el espacio climático están dados por la influencia en las variaciones de precipitación y temperatura de nuestra zona de estudio, provocando que los ecosistemas y, por ende, las especies que los conforman, modifiquen sus rangos de distribución tanto latitudinal como longitudinalmente. Se estima que, a futuro bajo el escenario en análisis, los ecosistemas presentes en la Zona Sur sufrirían un incremento tanto en los índices de precipitación como de temperatura, de 14,9 mm y 3,56 °C en promedio, respectivamente (tabla 1, Anexo 4 y 5). Si bien existe un incremento de precipitación, esto no ocurriría para todos los ecosistemas presentes en el área de investigación, de acuerdo a los resultados obtenidos los ecosistemas de los pisos bioclimáticos como Montano, Montano Alto y Montano alto superior, la precipitación decrecería en promedio 19,23 mm, 18,76 mm y 22,39 mm respectivamente; mientras que en los ecosistemas de pisos bioclimáticos como Tierras bajas, Piemontano y Montano bajo el incremento de precipitación media sería de 23,81 mm, 43,86 mm y 29 mm, respectivamente. Por otro lado, las variaciones de temperatura sufrirían un incremente para todos los ecosistemas presentes en la Zona Sur del Ecuador.
Por otro lado, es probable que las nuevas condiciones climáticas favorezcan el desarrollo de formaciones vegetales y, por consiguiente, la expansión o formación de otros ecosistemas. Sin embargo, dado que la distribución de las especies se encuentra limitado por varios factores, estos resultados se deberían interpretar desde el punto de vista de la idoneidad en las condiciones climáticas de las formaciones vegetales más no como modelos de distribución de ecosistemas. Así también se daría la posibilidad de algunos ecosistemas permanecer en tiempo y espacio sin presentar variaciones en su distribución.
Tabla 1. Variación en el espacio climático (temperatura y precipitación) actual y futuro (RCP6.0.-2080), en la Zona Sur del Ecuador.
|
Ecosistema |
Pisos Bioclimáticos |
Precipitación |
Temperatura |
||||
|
Actual |
RCP 6.0 |
Variación |
Actual |
RCP 6.0 |
Variación |
||
|
AdBn01 |
Montano bajo |
80,7 |
284,4 |
+ 203,7 |
20,06 |
23,39 |
+ 3,33 |
|
AmMn01 |
Montano |
107,7 |
125,0 |
+ 17,3 |
17,05 |
19,81 |
+ 2,76 |
|
AsAn01 |
Montano alto |
134,1 |
125,7 |
- 8,4 |
10,75 |
14,93 |
+ 4,18 |
|
AsMa01 |
Montano |
191,4 |
159,4 |
- 32,0 |
11,62 |
17,01 |
+ 5,38 |
|
AsMn02 |
Montano |
107,4 |
99,7 |
- 7,7 |
13,57 |
15,19 |
+ 1,62 |
|
AsSn01 |
Montano alto superior |
149,5 |
124,3 |
- 25,2 |
9,47 |
13,06 |
+ 3,59 |
|
BdBn01 |
Montano bajo |
132,7 |
153,5 |
+ 20,8 |
17,31 |
20,99 |
+ 3,68 |
|
BdPn01 |
Piemontano |
78,7 |
123,6 |
+ 44,9 |
22,64 |
25,13 |
+ 2,50 |
|
BdTc01 |
Tierras bajas |
59,8 |
104,6 |
+ 44,8 |
24,48 |
26,62 |
+ 2,14 |
|
BdTc02 |
Tierras bajas |
64,5 |
92,4 |
+ 28,0 |
24,52 |
26,80 |
+ 2,28 |
|
BeBn01 |
Montano bajo |
126,2 |
153,5 |
+ 27,3 |
16,37 |
20,51 |
+ 4,13 |
|
BePn01 |
Piemontano |
183,9 |
145,2 |
- 38,6 |
20,92 |
24,63 |
+ 3,71 |
|
BePn02 |
Piemontano |
133,8 |
176,4 |
+ 42,6 |
21,09 |
24,88 |
+ 3,79 |
|
BeTc02 |
Tierras bajas |
113,3 |
142,8 |
+ 29,5 |
23,41 |
26,57 |
+ 3,17 |
|
BmBn01 |
Montano bajo |
119,0 |
145,9 |
+ 26,9 |
17,20 |
20,58 |
+ 3,39 |
|
BmBn02 |
Montano bajo |
83,5 |
108,1 |
+ 24,6 |
20,20 |
23,19 |
+ 2,99 |
|
BmPn01 |
Piemontano |
106,9 |
145,1 |
+ 38,2 |
21,07 |
24,48 |
+ 3,40 |
|
BmPn02 |
Piemontano |
108,3 |
162,5 |
+ 54,2 |
21,19 |
24,40 |
+ 3,22 |
|
BmTc01 |
Tierras bajas |
122,8 |
131,4 |
+ 8,6 |
23,70 |
26,59 |
+ 2,89 |
|
BsAn02 |
Montano alto |
142,4 |
119,7 |
- 22,7 |
10,42 |
13,61 |
+ 3,20 |
|
BsAn04 |
Montano alto |
148,1 |
123,0 |
- 25,1 |
10,66 |
13,62 |
+ 2,96 |
|
BsBa02 |
Montano bajo |
201,8 |
245,6 |
+ 43,8 |
17,32 |
21,92 |
+ 4,60 |
|
BsBa03 |
Montano bajo |
210,0 |
261,6 |
+ 51,5 |
17,22 |
21,96 |
+ 4,74 |
|
BsBn02 |
Montano bajo |
142,8 |
171,4 |
+ 28,6 |
16,24 |
20,46 |
+ 4,22 |
|
BsBn04 |
Montano bajo |
219,1 |
126,9 |
- 92,2 |
16,95 |
20,66 |
+ 3,71 |
|
BsBn05 |
Montano bajo |
183,2 |
138,0 |
- 45,1 |
16,67 |
20,67 |
+ 4,01 |
|
BsMa01 |
Montano |
228,9 |
214,5 |
- 14,4 |
14,10 |
19,60 |
+ 5,50 |
|
BsMa02 |
Montano |
191,5 |
183,8 |
- 7,7 |
14,52 |
19,47 |
+ 4,95 |
|
BsMn02 |
Montano |
147,0 |
136,1 |
- 10,9 |
13,05 |
16,64 |
+ 3,58 |
|
BsMn03 |
Montano |
225,8 |
124,2 |
- 101,6 |
9,98 |
14,70 |
+ 4,72 |
|
BsMn04 |
Montano |
134,2 |
137,4 |
+ 3,2 |
13,57 |
16,70 |
+ 3,13 |
|
BsPa02 |
Piemontano |
166,9 |
246,5 |
+ 79,7 |
20,36 |
23,81 |
+ 3,44 |
|
BsPa03 |
Piemontano |
209,3 |
247,4 |
+ 38,1 |
19,99 |
23,76 |
+ 3,77 |
|
BsPa04 |
Piemontano |
184,2 |
258,9 |
+ 74,7 |
19,98 |
23,71 |
+ 3,74 |
|
BsPn02 |
Piemontano |
168,1 |
196,8 |
+ 28,7 |
19,34 |
24,19 |
+ 4,85 |
|
BsPn04 |
Piemontano |
122,7 |
198,8 |
+ 76,1 |
19,66 |
23,30 |
+ 3,65 |
|
BsTc05 |
Tierras bajas |
80,9 |
94,5 |
+ 13,6 |
24,08 |
27,20 |
+ 3,12 |
|
HsSn02 |
Montano alto superior |
129,5 |
113,0 |
- 16,5 |
10,52 |
12,97 |
+ 2,45 |
|
HsSn04 |
Montano alto superior |
146,6 |
121,1 |
- 25,5 |
9,45 |
12,78 |
+ 3,33 |
|
HsTc03 |
Tierras bajas |
78,5 |
96,9 |
+ 18,4 |
24,75 |
27,46 |
+ 2,71 |
Fuente: resultados del proceso de modelización. Elaboración propia.
El tipo y número de variables que se emplearon para modelar la distribución de los ecosistemas en el espacio geográfico, dependen estrictamente de las características de cada uno de los ecosistemas y de la escala de trabajo en la que se realice la modelización (Sainz, 2019).
La frecuencia de uso de las variables topográficas es respaldado por la tesis central de ecología vegetal mencionada por Brovkin (2002), en donde las condiciones climáticas ejercen control dominante sobre la distribución espacial de los principales tipos de vegetación a escala global, mientras que a escala de paisaje (como la analizada en esta investigación), la contribución de factores secundarios como el tipo de suelo y topografía también son importantes, influyendo en factores como temperatura, radiación solar, hidrología, entre otros. Estas variables cobran aún más relevancia en su uso al tratarse de una provincia que se sitúa en una de las zonas más irregulares del país, con predominancia montañosa, de llanuras raras, pequeñas y profundas y con un clima inestable (Alvarado & Benavidez-Silva, 2022).
Si bien las áreas de recambio nos indican una idoneidad en las condiciones climáticas para el establecimiento de los ecosistemas, sería impreciso indicar que ecosistemas llegarían a ocupar estas zonas, generando cierta incertidumbre (Marquet et al., 2010) debido a que los ecosistemas están compuestos por especies vegetales, componentes estructurales que controlan numerosos procesos ecológicos y sustentan una amplia variedad de organismos (Ramachandran et al., 2020) , por lo tanto, es necesario hipotetizar si esas condiciones climáticas son suficientes como para que las principales formaciones vegetales que dominan los ecosistemas se puedan desarrollar; porque las condiciones climáticas están presentes, pero sería incierto indicar si las formaciones vegetales tendrán la capacidad de colonizar estas zonas geográficas o si las especies que habitan estas zonas serían capaces de adaptarse a los cambios de las variables bioclimáticas en el futuro. Este comportamiento difuso de los modelos, se debe a la relación que existe entre espacio geográfico con el espacio climático, es decir que, “para cada punto en el espacio geográfico existe solo un punto en el espacio ambiental, pero para cada punto en el espacio ambiental se puede encontrar varios puntos del espacio geográfico” (Soberón et al., 2017), lo que eventualmente generaría que los ecosistemas potencialmente podrían compartir un espacio climático-geográfico similar y una posible sobre posición de condiciones climáticas para el establecimiento de diferentes ecosistemas en el futuro.
Estos cambios en el espacio climático están dados por la influencia en las variaciones de precipitación y temperatura, provocando que las condiciones climáticas que hoy caracterizan los diferentes ecosistemas sean diferentes en el futuro, y por lo tanto, la vegetación primaria que conforman los diferentes ecosistemas tienda a modificar sus rangos de distribución tanto latitudinal como longitudinalmente, en busca de áreas que presenten las condiciones climáticas y topográficas adecuadas para su desarrollo y con ello la conformación de los diferentes ecosistemas (Benavidez-Silva et al., 2021; Pliscoff, 2022).
Estos incrementos son respaldados por las proyecciones realizadas por el Panel Intergubernamental de Expertos sobre el Cambio Climático (IPCC, 2014), quienes estiman un incremento de la temperatura entre 1,4 a 3,1 °C con una probabilidad >66%, bajo el mismo escenario en análisis. Así mismo, los resultados encontrados concuerdan con las proyecciones realizadas de precipitación y temperatura para el Ecuador continental por Armenta et al. (2016), para un mismo escenario y el mismo horizonte temporal, se estima que la temperatura podría incrementarse de 1,5 a 3,5 °C, y la precipitación sufriría un incremento de hasta 20 mm para la Zona Sur del Ecuador. Y, con las proyecciones realizadas por Aguirre et al. (2015) para la Zona Sur del Ecuador, para los dos escenarios, un optimista (RCP 2.6) y un pesimista (RCP 8.5) y para el año 2050, en donde la precipitación se incrementaría hasta el 17 y 31% respectivamente, concentrándose los mayores cambios en la provincia del el Oro; mientras que la temperatura en promedio se incrementaría hasta 1,56 °C para un el escenario RCP 2.6 y un aumento de 2,37 °C para el escenario RCP 8.5, concentrándose los mayores aumentos en la provincia de Zamora.
En cuanto a las proyecciones del modelo para el 2080, se esperan cambios futuros para la mayoría de los ecosistemas. Los modelos sugieren una posible disminución de idoneidad climática, especialmente para los ecosistemas de Bosque montano alto, Bosque montano, Bosque montano alto y Páramo. Una posible explicación estaría asociado con la disminución de precipitación en los meses más secos (Bio 18), volviéndose más sensibles a la variabilidad climática, principalmente debido a la disminución de la disponibilidad de humedad (Bio 15) (Anjos & de Toledo, 2018). Por lo tanto, los bosques se volverían más vulnerables a las sequias (Rocha, 2022).
El cambio climático es una amenaza global que afecta el equilibrio del sistema climático del planeta y tiene graves consecuencias para la salud humana y la biodiversidad. El aumento de la temperatura y los cambios en los patrones de precipitación causados por el cambio climático impactan negativamente los ecosistemas terrestres, obligando a las especies a modificar sus límites geográficos y llevando a la redistribución de la vida en el planeta. Algunos organismos pueden evolucionar para adaptarse a estos cambios, mientras que otros pueden desaparecer. Los ecosistemas en la Zona Sur de Ecuador, que incluyen las provincias de Loja, Zamora Chinchipe y El Oro, son vulnerables a los efectos del cambio climático debido a la fragmentación histórica causada por cambios en el uso de la tierra, como el crecimiento urbano, la deforestación, la minería y la expansión de la frontera agrícola. Los modelos correlativos se pueden utilizar para predecir la distribución potencial de los ecosistemas en la Zona Sur de Ecuador hacia el año 2080 bajo un escenario de cambio climático, lo que puede ayudar a comprender las variaciones espaciales y temporales que experimentan los ecosistemas en respuesta a diferentes escenarios climáticos futuros. Esta información puede ser utilizada para gestionar y proteger los ecosistemas mediante la creación de nuevas áreas de conservación. La identificación de las principales variables biofísicas que determinan la distribución potencial de los ecosistemas puede ayudar a predecir la distribución futura de los ecosistemas y su respuesta al cambio climático.
Los autores agradecen al proyecto Biogeografía del cambio climático en el Sur del Ecuador: Dinámicas de cambio de uso de suelo y su influencia en la distribución espacial y temporal de especies vegetales (20 – DI – FARNR - 2021), a la Dirección de Investigación por el financiamiento del estudio y a la Carrera de Ingeniería Ambiental de la Universidad Nacional de Loja.
Todas/os las/os autoras/es se comprometen a comunicar cualquier conflicto de intereses existente o potencial con relación a la publicación de su artículo.
La contribución de cada autor al presente artículo ha sido la siguiente:
Aguirre Mendoza, Z., Aguirre Mendoza, N., & Muñoz, J. (2017). Biodiversidad de la provincia de Loja, Ecuador. Arnaldoa, 24(2), 523–542. https://doi.org/10.22497/arnaldoa.242.24206
Aguirre, N., Eguiguren, P., Maita, J., Coronel, V., Samaniego, N., Ojeda, T., & Aguirre, Z. (2015). Vulnerabilidad al cambio climático en la Región Sur del Ecuador: Potenciales impactos en los ecosistemas, producción de biomasa y producción hídrica. Universidad Nacional de Loja y Servicio Forestal de los Estados Unidos.
Allouche, O., Tsoar, A., & Kadmon, R. (2006). Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). Journal of Applied Ecology, 43(6), 1223–1232. https://doi.org/10.1111/j.1365-2664.2006.01214.x
Alvarado, M., & Benavidez-Silva, C. (2022). Cambio y continuidad en la valorización del espacio agrario de Loja. SIPAE. https://doi.org/10.13140/RG.2.2.11054.38725
Anjos, L. J. S., & de Toledo, P. M. (2018). Measuring resilience and assessing vulnerability of terrestrial ecosystems to climate change in South America. PLOS ONE, 13(3), e0194654. https://doi.org/10.1371/journal.pone.0194654
Armenta, G., Villa, J., & Jácome, P. (2016). Proyecciones climáticas de precipitación y temperatura para Ecuador, bajo distintos escenarios de cambio climático. PNUD. https://info.undp.org/docs/pdc/Documents/ECU/14%20Proyecciones%20de%20Clima%20Futuro%20para%20Ecuador%20en%20base%20a%20IPCC-AR5.pdf
Ashraf, U., Peterson, A. T., Chaudhry, M. N., Ashraf, I., Saqib, Z., Ahmad, S. R., & Ali, H. (2017). Ecological niche model comparison under different climate scenarios: A case study of Olea spp. in Asia. Ecosphere, 8(5), 1–13. https://doi.org/10.1002/ecs2.1825
Atauchi, P. J., Aucca-Chutas, C., Ferro, G., & Prieto-Torres, D. A. (2020). Present and future potential distribution of the endangered Anairetes alpinus (Passeriformes: Tyrannidae) under global climate change scenarios. Journal of Ornithology, 161(3), 723–738. https://doi.org/10.1007/s10336-020-01762-z
Benavidez-Silva, C., Jensen, M., & Pliscoff, P. (2021). Future scenarios for land use in chile: Identifying drivers of change and impacts over protected area system. Land, 10(4), 1 – 21. https://doi.org/10.3390/land10040408
Brehm, G., Homeier, J., Fiedler, K., Kottke, I., Illig, J., Nöske, N. M., Werner, F. A., & Breckle, S. W. (2008). Mountain Rain Forests in Southern Ecuador as a Hotspot of Biodiversity – Limited Knowledge and Diverging Patterns. Gradients in a Tropical Mountain Ecosystem of Ecuador. Ecological Studies, 198, 15–23. https://doi.org/10.1007/978-3-540-73526-7_2
Brovkin, V. (2002). Climate-vegetation interaction. Journal de Physique IV (Proceedings), 12(10), 57–72. https://doi.org/10.1051/jp4:20020452
Brun, P., Thuiller, W., Chauvier, Y., Pellissier, L., Wüest, R. O., Wang, Z., & Zimmermann, N. E. (2020). Model complexity affects species distribution projections under climate change. Journal of Biogeography, 47(1), 130–142. https://doi.org/10.1111/jbi.13734
Busch, J., Dave, R., Hannah, L., Cameron, A., Rasolohery, A., Roehrdanz, P., & Schatz, G. (2012). Climate Change and the Cost of Conserving Species in Madagascar. Conservation Biology, 26(3), 408–419. https://doi.org/10.1111/j.1523-1739.2012.01838.x
Carroll, C., Roberts, D. R., Michalak, J. L., Lawler, J. J., Nielsen, S. E., Stralberg, D., Hamann, A., Mcrae, B. H., & Wang, T. (2017). Scale‐dependent complementarity of climatic velocity and environmental diversity for identifying priority areas for conservation under climate change. Global Change Biology, 23(11), 4508–4520. https://doi.org/10.1111/gcb.13679
CEPAL. (2017). Síntesis de políticas públicas sobre cambio climático. El cambio climático y sus efectos en la biodiversidad de América Latina. CEPAL. https://repositorio.cepal.org/handle/11362/45692
Colwell, R. K., Brehm, G., Cardelús, C. L., Gilman, A. C., & Longino, J. T. (2008). Global warming, elevational range shifts, and lowland biotic attrition in the Wet Tropics. Science, 322, 258–261. http://www.sciencemag.org/cgi/doi/10.1126/science.1162547
Correa-Lima, A. P. A., Varassin, I. G., Barve, N., & Zwiener, V. P. (2019). Spatiotemporal effects of climate change on the geographical distribution and flowering phenology of hummingbird-pollinated plants. Annals of Botany, 124(3), 389–398. https://doi.org/10.1093/aob/mcz079
Cueva, J. L. (2012). Elaboración y análisis del estado de la cobertura vegetal de la provincia de Loja - Ecuador. [Tesis de fin de máster, Universidad Internacional de Andalucía]. Repositorio Institucional. http://hdl.handle.net/10334/1794
Dagtekin, D., Şahan, E. A., Denk, T., Köse, N., & Dalfes, H. N. (2020). Past, present and future distributions of Oriental beech (Fagus orientalis) under climate change projections. PLoS ONE, 15, 1–19. https://doi.org/10.1371/journal.pone.0242280
De Pando, B. B., & De Giles, J. P. (2007). Aplicación de modelos de distribución de especies a la conservación de la biodiversidad en el sureste de la Península Ibérica. GeoFocus, 7, 100–119. http://geofocus.rediris.es/2007/Articulo6_2007.pdf
Farr, T. G., Rosen, P. A., Caro, E., Crippen, R., Duren, R., Hensley, S., Kobrick, M., Paller, M., Rodriguez, E., Roth, L., Seal, D., Shaffer, S., Shimada, J., Umland, J., Werner, M., Oskin, M., Burbank, D., & Alsdorf, D. (2007). The Shuttle Radar Topography Mission. Reviews of Geophysics, 45(2), 880–881. https://doi.org/10.1029/2005RG000183
Fletcher, R., & Fortin, M.-J. (2018). Spatial Ecology and Conservation Modeling: Applications with R. SPRINGER. https://doi.org/10.1007/978-3-030-01989-1
Gaglio, M., Aschonitis, V., Pieretti, L., Santos, L., Gissi, E., Castaldelli, G., & Fano, E. A. (2019). Modelling past, present and future Ecosystem Services supply in a protected floodplain under land use and climate changes. Ecological Modelling, 403, 23–34. https://doi.org/10.1016/j.ecolmodel.2019.04.019
Gang, C., Zhang, Y., Wang, Z., Chen, Y., Yang, Y., Li, J., Cheng, J., Qi, J., & Odeh, I. (2017). Modeling the dynamics of distribution, extent, and NPP of global terrestrial ecosystems in response to future climate change. Global and Planetary Change, 148, 153–165. https://doi.org/10.1016/j.gloplacha.2016.12.007
Garzón-Santomaro, C., Prieto-Albuja, F., Brito. J., & Mena-Jaén, J. (Eds.). (2019). Propuesta para el establecimiento del Subsistema de Áreas Naturales de Conservación y Diseño del Corredor Ecológico de la provincia de El Oro: Una guía para el desarrollo de estrategias del investigación, conservación y manejo de la biodiversidad orense. GADPEO – INABIO.
Guisan, A., Zimmermann, N. E. (2000). Predictive habitat distribution models in ecology. Ecological Modelling, 135, 147–186. https://doi.org/10.1016/S0304-3800(00)00354-9
Hao, T., Elith, J., Guillera-Arroita, G., & Lahoz-Monfort, J. J. (2019). A review of evidence about use and performance of species distribution modelling ensembles like BIOMOD. Diversity and Distributions, 25(5), 839–852. https://doi.org/10.1111/ddi.12892
Harrison, P. Berry, P., Dawson, T. (2003). Modelling Natural Resource Responses to Climate Change (the MONARCH project): an introduction. Journal for Nature Conservation, 11(1), 3 – 4. https://doi.org/10.1078/1617-1381-00032.
Hylander, K., Greiser, C., Christiansen, D. M., & Koelemeijer, I. A. (2022). Climate adaptation of biodiversity conservation in managed forest landscapes. Conservation Biology, 36(3), 1–9. https://doi.org/10.1111/cobi.13847
IPCC. (2008). Cambio climático. In Grupo Intergubernamental de Expertos sobre el Cambio Climático. IPCC. https://www.ipcc.ch/site/assets/uploads/2018/02/ar4_syr_sp.pdf
IPCC. (2014). Cambio Climático 2014: Informe de síntesis. Contribución de los Grupos de trabajo I, II y III al Quinto Informe de Evaluación del Grupo Intergubernamental de Expertos Sobre el Cambio Climático. https://www.ipcc.ch/site/assets/uploads/2018/02/SYR_AR5_FINAL_full_es.pdf
Kadmon, R., Farber, O., & Danin, A. (2003). A systematic analysis of factors affecting the performance of climatic envelope models. Ecological Applications, 13(3), 853–867. https://doi.org/10.1890/1051-0761(2003)013[0853:ASAOFA]2.0.CO;2
Lozano, P. (2002). Los tipos de bosque en el sur del Ecuador. In Z. Aguirre, J.E. Madsen, E., Cotton & H. Balslev (Eds.), Botánica Austroecuatoriana. Estudios sobre los recursos vegetales en las provincial de El Oro, Loja y Zamora-Chinchipe. Abya-Yala, Quito (pp. 29–49). https://www.researchgate.net/publication/233936622
MAE. (2013). Sistema de clasificación de los ecosistemas de Ecuador Continental. In Subsecretaría de Patrimonio Natural. Quito. http://ide.ambiente.gob.ec/mapainteractivo/
Malhi, Y., Franklin, J., Seddon, N., Solan, M., Turner, M. G., Field, C. B., & Knowlton, N. (2020). Climate change and ecosystems: Threats, opportunities and solutions. Philosophical Transactions of the Royal Society. Biological Sciences, 375(1794), 1–8. https://doi.org/10.1098/rstb.2019.0104
Manchego, C. E., Hildebrandt, P., Cueva, J., Espinosa, C. I., Stimm, B., & Günter, S. (2017). Climate change versus deforestation: Implications for tree species distribution in the dry forests of southern Ecuador. PLoS ONE, 12(12), 1–19. https://doi.org/10.1371/journal.pone.0190092
Marquet, P., Abades, S., Armesto, J., Barria, I., Arroyo, M., Cavieres, L., Gajardo, R., Garin, C., Labra, F., Meza, F., Prado, C., Ramirez, P., & Vicuña, S. (2010). Estudio de vulnerabilidad de la biodiversidad terrestre en la Eco-región Mediterránea, a nivel de ecosistemas y especies, y medidas de adaptación frente a escenarios de cambio climático. CIREN. https://bibliotecadigital.ciren.cl/handle/20.500.13082/26033
Parry, L., Radel, C., Adamo, S. B., Clark, N., Counterman, M., Flores-Yeffal, N., Pons, D., Romero-Lankao, P., & Vargo, J. (2019). The (in)visible health risks of climate change. Social Science and Medicine, 241, 1–15. https://doi.org/10.1016/j.socscimed.2019.112448
Pecl, G. T., Araújo, M. B., Bell, J. D., Blanchard, J., Bonebrake, T. C., Chen, I. C., Clark, T. D., Colwell, R. K., Danielsen, F., Evengård, B., Falconi, L., Ferrier, S., Frusher, S., Garcia, R. A., Griffis, R. B., Hobday, A. J., Janion-Scheepers, C., Jarzyna, M. A., Jennings, S., … Williams, S. E. (2017). Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science, 355, 1–9. https://doi.org/10.1126/science.aai9214
Pliscoff, P. (2022). Actualización de las áreas protegidas de Chile: análisis de representatividad y riesgo climático. Centro de estudios público (CEP). https://www.cepchile.cl/investigacion/actualizacion-de-las-areas-protegidas-de-chile-analisis-de/
Pliscoff, P., & Fuentes-Castillo, T. (2011). Modelación de la distribución de especies y ecosistemas en el tiempo y en el espacio: Una revisión de las nuevas herramientas y enfoques disponibles. Revista de Geografia Norte Grande, 2011(48), 61–79. https://doi.org/10.4067/s0718-34022011000100005
Quesada-Quirós, M., Acosta-Vargas, L. G., Arias-Aguilar, D., & Rodríguez-González, A. (2016). Modelación de nichos ecológicos basado en tres escenarios de cambio climático para cinco especies de plantas en zonas altas de Costa Rica. Revista Forestal Mesoamericana Kurú, 14(34), 01–12. https://doi.org/10.18845/rfmk.v14i34.2991
Ramachandran, R. M., Roy, P. S., Chakravarthi, V., Joshi, P. K., & Sanjay, J. (2020). Land use and climate change impacts on distribution of plant species of conservation value in Eastern Ghats, India: a simulation study. Environmental Monitoring and Assessment, 192(2), 1–22. https://doi.org/10.1007/s10661-019-8044-5
Ren, Z., Peng, H., & Liu, Z. W. (2016). The rapid climate change-caused dichotomy on subtropical evergreen broad-leaved forest in Yunnan: Reduction in habitat diversity and increase in species diversity. Plant Diversity, 38(3), 142–148. https://doi.org/10.1016/j.pld.2016.04.003
Rocha, J. C. (2022). Ecosystems are showing symptoms of resilience loss. Environmental Research Letters, 17(6), 1–13. https://doi.org/10.1088/1748-9326/ac73a8
Sainz Villegas, S. (2019). Efectos del cambio climático en la distribución geográfica de la especie invasora Asparagopsis armata Harvey, 1855. [Tesis fin de máster, Máster Universitario en Gestión Integrada de Sistemas Hídricos]. Universidad de Cantabria.
Samaniego-Rojas, N., Eguiguren, P., Maita, J., & Aguirre, N. (2015). Clima de la Región Sur el Ecuador: historia y tendencias. In Aguirre et al. (Eds). Cambio climático y Biodiversidad: estudio de caso de los páramos del Parque Nacional Podocarpus, Ecuador (pp. 43–63). Programa de biodiversidad y Servicios Ecosistémicos. Universidad Nacional de Loja, Ecuador.
Soberón, J., Osorio-Olvera, L., & Peterson, T. (2017). Diferencias conceptuales entre modelación de nichos y modelación de áreas de distribución. Revista Mexicana de Biodiversidad, 88(2), 437–441. https://doi.org/10.1016/j.rmb.2017.03.011
Tapia-Armijos, M. F., Homeier, J., Espinosa, C. I., Leuschner, C., & De La Cruz, M. (2015). Deforestation and forest fragmentation in south Ecuador since the 1970s - Losing a hotspot of biodiversity. PLoS ONE, 10(9), 1–18. https://doi.org/10.1371/journal.pone.0133701
Thomas, C. D. (2020). The development of Anthropocene biotas. Philosophical Transactions of the Royal Society. Biological Sciences, 375, 1–9. https://doi.org/10.1098/rstb.2019.0113
Thuiller, W. (2003). BIOMOD - optimizing predictions of species distributions and projecting potential future shifts under global change. Global Change Biology, 20(12), 3591–3592. https://doi.org/10.1111/gcb.12728
Thuiller, W., Georges, D., Gueguen, M., Engler, R., Breiner, F., & Lafourcade, B. (2022). biomod2: Ensemble Platform for Species Distribution Modeling. R Package Version 4.0.
Vazquez, M. A., Freire, J. F., & Suárez, L. (2005). Biodiversidad en los bosques secos de la zona de Cerro Negro-Cazaderos, occidente de la provincia de Loja: Un reporte de las evaluaciones ecológicas y socioeconómicas rápidas. In EcoCiencia, MAE y Proyecto Bosque Seco. Quito.
Weiskopf, S. R., Rubenstein, M. A., Crozier, L. G., Gaichas, S., Griffis, R., Halofsky, J. E., Hyde, K. J. W., Morelli, T. L., Morisette, J. T., Muñoz, R. C., Pershing, A. J., Peterson, D. L., Poudel, R., Staudinger, M. D., Sutton-Grier, A. E., Thompson, L., Vose, J., Weltzin, J. F., & Whyte, K. P. (2020). Climate change effects on biodiversity, ecosystems, ecosystem services, and natural resource management in the United States. Science of the Total Environment, 733, 1–18. https://doi.org/10.1016/j.scitotenv.2020.137782
Tabla síntesis con resultado de la evaluación de todos los modelos para todos los ecosistemas modelados (el símbolo – significa ausencia de evaluación para la combinación específica de ecosistema y modelo).
|
CÓDIGO ECOSISTEMA |
MÉTRICA DE EVALUACIÓN |
MODELOS |
|||||||||
|
GLM |
GBM |
GAM |
CTA |
ANN |
SRE |
FDA |
MARS |
RF |
MAXENT |
||
|
AdBn01 |
TSS |
0,99 |
1,00 |
0,92 |
0,93 |
0,92 |
0,86 |
1,00 |
1,00 |
1,00 |
1,00 |
|
ROC |
1,00 |
1,00 |
0,96 |
0,93 |
0,96 |
0,93 |
1,00 |
1,00 |
1,00 |
1,00 |
|
|
AmMn01 |
TSS |
0,91 |
0,91 |
0,89 |
0,82 |
0,87 |
0,60 |
0,77 |
0,93 |
0,93 |
0,83 |
|
ROC |
0,97 |
0,98 |
0,96 |
0,93 |
0,95 |
0,80 |
0,94 |
0,98 |
0,99 |
0,98 |
|
|
AsAn01 |
TSS |
1,00 |
1,00 |
1,00 |
0,97 |
0,89 |
0,90 |
0,96 |
1,00 |
1,00 |
0,99 |
|
ROC |
1,00 |
1,00 |
1,00 |
0,99 |
0,94 |
0,95 |
0,98 |
1,00 |
1,00 |
1,00 |
|
|
AsMa01 |
TSS |
0,99 |
0,99 |
0,77 |
0,98 |
0,85 |
0,66 |
- |
0,97 |
0,99 |
0,94 |
|
ROC |
1,00 |
1,00 |
0,89 |
0,99 |
0,93 |
0,83 |
- |
0,99 |
1,00 |
0,99 |
|
|
AsMn02 |
TSS |
0,96 |
0,96 |
0,89 |
0,93 |
- |
0,61 |
0,94 |
0,95 |
0,96 |
0,86 |
|
ROC |
0,98 |
0,98 |
0,95 |
0,97 |
- |
0,80 |
0,99 |
0,98 |
0,98 |
0,93 |
|
|
AsSn01 |
TSS |
0,91 |
0,89 |
- |
0,87 |
0,85 |
0,60 |
0,94 |
0,91 |
0,87 |
0,77 |
|
ROC |
0,99 |
0,96 |
- |
0,94 |
0,93 |
0,80 |
0,99 |
0,99 |
0,97 |
0,89 |
|
|
BdBn01 |
TSS |
0,97 |
0,99 |
0,96 |
0,98 |
- |
0,79 |
0,95 |
0,97 |
0,99 |
0,99 |
|
ROC |
0,99 |
1,00 |
0,98 |
0,99 |
- |
0,90 |
0,99 |
0,99 |
1,00 |
1,00 |
|
|
BdPn01 |
TSS |
0,95 |
0,84 |
0,96 |
0,85 |
- |
0,51 |
0,85 |
0,95 |
0,85 |
0,94 |
|
ROC |
0,99 |
0,95 |
0,99 |
0,94 |
- |
0,76 |
0,96 |
0,99 |
0,95 |
0,99 |
|
|
BdTc01 |
TSS |
0,98 |
0,96 |
0,70 |
0,81 |
0,82 |
0,70 |
0,97 |
0,99 |
0,97 |
0,99 |
|
ROC |
0,99 |
0,99 |
0,85 |
0,91 |
0,91 |
0,85 |
0,98 |
0,99 |
0,99 |
1,00 |
|
|
BdTc02 |
TSS |
0,98 |
0,97 |
0,88 |
0,93 |
- |
0,54 |
0,96 |
0,98 |
0,94 |
0,97 |
|
ROC |
1,00 |
0,99 |
0,94 |
0,97 |
- |
0,77 |
0,98 |
1,00 |
0,99 |
1,00 |
|
|
BeBn01 |
TSS |
0,93 |
0,98 |
0,84 |
0,94 |
0,77 |
0,72 |
0,85 |
0,80 |
0,94 |
0,89 |
|
ROC |
0,99 |
0,99 |
0,92 |
0,97 |
0,92 |
0,86 |
0,96 |
0,90 |
0,99 |
0,96 |
|
|
BePn01 |
TSS |
1,00 |
1,00 |
1,00 |
0,94 |
0,92 |
0,65 |
0,96 |
0,94 |
1,00 |
0,70 |
|
ROC |
1,00 |
1,00 |
1,00 |
0,97 |
0,97 |
0,82 |
0,99 |
0,97 |
1,00 |
0,85 |
|
|
BePn02 |
TSS |
0,89 |
0,94 |
0,63 |
0,72 |
0,85 |
0,56 |
0,84 |
0,92 |
0,93 |
0,85 |
|
ROC |
0,98 |
0,99 |
0,82 |
0,86 |
0,92 |
0,78 |
0,97 |
0,98 |
0,99 |
0,96 |
|
|
BeTc02 |
TSS |
0,99 |
1,00 |
0,80 |
0,99 |
0,97 |
0,70 |
0,98 |
0,99 |
1,00 |
0,97 |
|
ROC |
1,00 |
1,00 |
0,90 |
1,00 |
0,98 |
0,85 |
0,99 |
1,00 |
1,00 |
1,00 |
|
|
BmBn01 |
TSS |
0,94 |
0,93 |
0,91 |
0,91 |
0,79 |
0,75 |
0,82 |
0,91 |
0,94 |
0,85 |
|
ROC |
0,99 |
0,99 |
0,98 |
0,94 |
0,90 |
0,87 |
0,97 |
0,97 |
0,99 |
0,96 |
|
|
BmBn02 |
TSS |
0,99 |
0,98 |
0,80 |
0,89 |
0,48 |
0,50 |
0,93 |
0,98 |
0,97 |
0,60 |
|
ROC |
1,00 |
1,00 |
0,90 |
0,94 |
0,74 |
0,75 |
0,99 |
1,00 |
1,00 |
0,80 |
|
|
BmPn01 |
TSS |
0,95 |
0,88 |
0,82 |
0,85 |
0,81 |
0,67 |
0,93 |
0,97 |
0,88 |
0,91 |
|
ROC |
0,99 |
0,98 |
0,92 |
0,91 |
0,97 |
0,84 |
0,99 |
0,99 |
0,99 |
0,97 |
|
|
BmPn02 |
TSS |
0,91 |
0,99 |
0,98 |
0,76 |
0,69 |
0,55 |
1,00 |
0,99 |
1,00 |
0,98 |
|
ROC |
0,92 |
1,00 |
0,99 |
0,88 |
0,84 |
0,78 |
1,00 |
1,00 |
1,00 |
1,00 |
|
|
BmTc01 |
TSS |
0,99 |
0,94 |
0,90 |
0,89 |
0,98 |
0,70 |
0,97 |
1,00 |
0,99 |
0,95 |
|
ROC |
1,00 |
1,00 |
0,95 |
0,95 |
0,99 |
0,85 |
1,00 |
1,00 |
1,00 |
1,00 |
|
|
BsAn02 |
TSS |
0,86 |
0,83 |
0,78 |
0,77 |
0,76 |
0,59 |
0,83 |
0,83 |
0,85 |
0,84 |
|
ROC |
0,95 |
0,97 |
0,90 |
0,92 |
0,92 |
0,79 |
0,93 |
0,93 |
0,97 |
0,97 |
|
|
BsAn04 |
TSS |
0,99 |
0,99 |
0,94 |
0,98 |
0,98 |
0,84 |
0,97 |
0,99 |
0,99 |
0,83 |
|
ROC |
1,00 |
1,00 |
0,97 |
0,99 |
0,99 |
0,92 |
0,99 |
1,00 |
1,00 |
0,92 |
|
|
BsBa02 |
TSS |
0,97 |
0,98 |
0,66 |
0,94 |
0,90 |
0,68 |
- |
0,44 |
0,89 |
0,98 |
|
ROC |
1,00 |
1,00 |
0,83 |
0,97 |
0,95 |
0,84 |
- |
0,51 |
0,99 |
1,00 |
|
|
BsBa03 |
TSS |
0,45 |
0,97 |
0,99 |
0,90 |
0,62 |
0,74 |
0,93 |
0,97 |
0,97 |
0,96 |
|
ROC |
0,78 |
0,99 |
0,99 |
0,95 |
0,80 |
0,87 |
0,97 |
0,99 |
0,99 |
0,99 |
|
|
BsBn02 |
TSS |
0,88 |
0,90 |
0,91 |
0,81 |
0,71 |
0,75 |
0,82 |
0,86 |
0,87 |
0,89 |
|
ROC |
0,98 |
0,97 |
0,98 |
0,92 |
0,93 |
0,88 |
0,96 |
0,98 |
0,98 |
0,98 |
|
|
BsBn04 |
TSS |
0,98 |
0,99 |
0,69 |
0,88 |
0,78 |
0,69 |
0,97 |
0,98 |
0,98 |
0,98 |
|
ROC |
1,00 |
1,00 |
0,85 |
0,94 |
0,96 |
0,84 |
0,99 |
1,00 |
1,00 |
1,00 |
|
|
BsBn05 |
TSS |
0,92 |
0,92 |
0,80 |
0,84 |
0,60 |
0,63 |
0,80 |
0,89 |
0,93 |
0,84 |
|
ROC |
0,99 |
0,97 |
0,90 |
0,91 |
0,80 |
0,82 |
0,94 |
0,99 |
0,97 |
0,92 |
|
|
BsMa01 |
TSS |
0,90 |
0,85 |
0,71 |
0,79 |
0,62 |
0,56 |
0,92 |
0,97 |
1,00 |
0,97 |
|
ROC |
0,96 |
0,98 |
0,85 |
0,90 |
0,80 |
0,78 |
0,98 |
0,99 |
1,00 |
0,99 |
|
|
BsMa02 |
TSS |
0,75 |
0,90 |
0,77 |
0,85 |
0,65 |
0,57 |
- |
0,96 |
0,85 |
0,95 |
|
ROC |
0,81 |
0,98 |
0,89 |
0,92 |
0,89 |
0,79 |
- |
0,98 |
0,97 |
0,98 |
|
|
BsMn02 |
TSS |
0,88 |
0,91 |
0,89 |
0,88 |
0,76 |
0,51 |
0,83 |
0,88 |
0,88 |
0,80 |
|
ROC |
0,98 |
0,96 |
0,96 |
0,94 |
0,88 |
0,75 |
0,95 |
0,97 |
0,98 |
0,96 |
|
|
BsMn03 |
TSS |
0,98 |
0,98 |
0,77 |
0,92 |
0,49 |
- |
0,92 |
0,96 |
0,95 |
0,77 |
|
ROC |
0,99 |
0,99 |
0,89 |
0,96 |
0,75 |
- |
0,98 |
0,99 |
0,99 |
0,89 |
|
|
BsMn04 |
TSS |
0,96 |
0,95 |
0,89 |
0,92 |
0,90 |
0,38 |
0,85 |
0,91 |
0,96 |
0,79 |
|
ROC |
0,99 |
0,99 |
0,95 |
0,96 |
0,97 |
0,69 |
0,97 |
0,98 |
0,99 |
0,90 |
|
|
BsPa02 |
TSS |
0,88 |
0,92 |
0,77 |
0,87 |
0,86 |
0,73 |
0,84 |
0,91 |
0,96 |
0,91 |
|
ROC |
0,96 |
0,94 |
0,89 |
0,94 |
0,93 |
0,86 |
0,97 |
0,99 |
0,99 |
0,99 |
|
|
BsPa03 |
TSS |
0,95 |
0,98 |
0,83 |
0,89 |
- |
0,73 |
0,96 |
0,97 |
0,98 |
0,96 |
|
ROC |
0,98 |
0,99 |
0,91 |
0,94 |
- |
0,86 |
0,98 |
0,98 |
1,00 |
0,99 |
|
|
BsPa04 |
TSS |
0,96 |
0,87 |
0,73 |
0,87 |
0,68 |
0,65 |
- |
0,89 |
0,87 |
0,91 |
|
ROC |
0,98 |
0,94 |
0,86 |
0,93 |
0,84 |
0,83 |
- |
0,98 |
0,98 |
0,98 |
|
|
BsPn02 |
TSS |
1,00 |
0,90 |
1,00 |
0,89 |
0,84 |
0,80 |
0,98 |
0,99 |
0,93 |
1,00 |
|
ROC |
1,00 |
0,95 |
1,00 |
0,95 |
0,92 |
0,90 |
0,99 |
1,00 |
1,00 |
1,00 |
|
|
BsPn04 |
TSS |
0,92 |
0,95 |
0,68 |
0,85 |
0,43 |
0,74 |
0,96 |
0,95 |
0,98 |
0,89 |
|
ROC |
0,99 |
0,99 |
0,84 |
0,94 |
0,72 |
0,87 |
0,99 |
0,99 |
1,00 |
0,94 |
|
|
BsTc05 |
TSS |
1,00 |
1,00 |
0,33 |
1,00 |
1,00 |
0,67 |
- |
1,00 |
1,00 |
1,00 |
|
ROC |
1,00 |
1,00 |
0,67 |
1,00 |
1,00 |
0,83 |
- |
1,00 |
1,00 |
1,00 |
|
|
HsSn02 |
TSS |
0,94 |
0,89 |
0,89 |
0,90 |
0,87 |
0,73 |
0,89 |
0,88 |
0,89 |
0,84 |
|
ROC |
0,99 |
0,99 |
0,95 |
0,97 |
0,97 |
0,86 |
0,97 |
0,98 |
0,98 |
0,94 |
|
|
HsSn04 |
TSS |
1,00 |
1,00 |
0,89 |
1,00 |
0,98 |
0,78 |
0,98 |
0,98 |
1,00 |
0,99 |
|
ROC |
1,00 |
1,00 |
0,94 |
1,00 |
0,99 |
0,89 |
0,99 |
0,98 |
1,00 |
1,00 |
|
|
HsTc03 |
TSS |
0,69 |
1,00 |
1,00 |
1,00 |
0,94 |
0,80 |
0,99 |
1,00 |
1,00 |
1,00 |
|
ROC |
0,85 |
1,00 |
1,00 |
1,00 |
0,97 |
0,90 |
1,00 |
1,00 |
1,00 |
1,00 |
|
Fuente: resultados del proceso de modelización. Elaboración propia.
Ecosistemas de la Zona Sur del Ecuador con su código de identificación y su piso bioclimático.
|
Ecosistemas Zona Sur |
Código MAE |
Piso bioclimático |
|
Bosque deciduo montano bajo del Catamayo-Alamor |
Bdbn01 |
Montano bajo |
|
Bosque deciduo piemontano del Catamayo-Alamor |
Bdpn01 |
Piemontano |
|
Bosque semideciduo piemontano del sur de la cordillera oriental de los Andes |
Bmpn02 |
Piemontano |
|
Bosque siempreverde montano bajo del Catamayo-Alamor |
Bsbn05 |
Montano bajo |
|
Bosque siempreverde piemontano del Catamayo-Alamor |
Bspn02 |
Piemontano |
|
Arbustal desértico del sur de los valles |
Adbn01 |
Montano bajo |
|
Arbustal semideciduo del sur de los valles |
Ammn01 |
Montano |
|
Arbustal siempreverde y herbazal del páramo |
Assn01 |
Montano alto superior |
|
Bosque bajo y arbustal deciduo de tierras bajas del Jama-Zapotillo |
Bdtc02 |
Tierras bajas |
|
Bosque deciduo de tierras bajas del Jama-Zapotillo |
Bdtc01 |
Tierras bajas |
|
Bosque semideciduo montano bajo del Catamayo-Alamor |
Bmbn01 |
Montano bajo |
|
Bosque semideciduo piemontano del Catamayo-Alamor |
Bmpn01 |
Piemontano |
|
Bosque siempreverde estacional montano bajo del Catamayo-Alamor |
Bebn01 |
Montano bajo |
|
Bosque siempreverde estacional piemontano del Catamayo-Alamor |
Bepn02 |
Piemontano |
|
Bosque siempreverde montano alto del Catamayo-Alamor |
Bsan04 |
Montano alto |
|
Bosque siempreverde montano del Catamayo-Alamor |
Bsmn04 |
Montano |
|
Arbustal siempreverde montano alto del páramo del Sur |
Asan01 |
Montano alto |
|
Arbustal siempreverde montano del sur de los Andes |
Asmn02 |
Montano |
|
Arbustal siempreverde y herbazal montano de la cordillera del Cóndor |
Asma01 |
Montano |
|
Bosque siempreverde montano alto del sur de la cordillera oriental de los Andes |
Bsan02 |
Montano alto |
|
Bosque siempreverde montano bajo de las cordilleras del Cóndor-Kutukú |
Bsba02 |
Montano bajo |
|
Bosque siempreverde montano bajo del sur de la cordillera oriental de los Andes |
Bsbn02 |
Montano bajo |
|
Bosque siempreverde montano bajo sobre mesetas de arenisca de las cordilleras del Cóndor-Kutukú |
Bsba03 |
Montano bajo |
|
Bosque siempreverde montano de las cordilleras del Cóndor-Kutukú |
Bsma02 |
Montano |
|
Bosque siempreverde montano del sur de la cordillera oriental de los Andes |
Bsmn02 |
Montano |
|
Bosque siempreverde montano sobre mesetas de arenisca de la cordillera del Cóndor |
Bsma01 |
Montano |
|
Bosque siempreverde piemontano de las cordilleras del Cóndor-Kutukú |
Bspa02 |
Piemontano |
|
Bosque siempreverde piemontano del sur de la cordillera oriental de los Andes |
Bspn04 |
Piemontano |
|
Bosque siempreverde piemontano sobre afloramientos de roca caliza de las cordilleras amazónicas |
Bspa03 |
Piemontano |
|
Bosque siempreverde piemontano sobre mesetas de arenisca de las cordilleras del Cóndor-Kutukú |
Bspa04 |
Piemontano |
|
Bosque y arbustal semideciduo del sur de los valles |
Bmbn02 |
Montano bajo |
|
Herbazal del páramo |
Hssn02 |
Montano alto superior |
|
Herbazal inundable del páramo |
Hssn04 |
Montano alto superior |
|
Bosque semideciduo de tierras bajas del Jama-Zapotillo |
Bmtc01 |
Tierras bajas |
|
Bosque siempreverde estacional de tierras bajas del Jama-Zapotillo |
Betc02 |
Tierras bajas |
|
Bosque siempreverde estacional piemontano de la cordillera occidental de los Andes |
Bepn01 |
Piemontano |
|
Bosque siempreverde montano bajo de la cordillera occidental de los Andes |
Bsbn04 |
Montano bajo |
|
Bosque siempreverde montano de la cordillera occidental de los Andes |
Bsmn03 |
Montano |
|
Herbazal inundable ripario de tierras bajas del Jama-Zapotillo |
Hstc03 |
Tierras bajas |
|
Manglar del Jama-Zapotillo |
Bstc05 |
Tierras bajas |
Fuente: MAE, 2013. Elaboración propia.
Variables explicativas implementadas para la modelización individual de los ecosistemas de la Zona Sur del Ecuador.
|
Ecosistema (código MAE) |
BIO1 |
BIO2 |
BIO3 |
BIO4 |
BIO5 |
BIO6 |
BIO7 |
BIO8 |
BIO9 |
BIO10 |
BIO11 |
BIO12 |
BIO13 |
BIO14 |
BIO15 |
BIO16 |
BIO17 |
BIO18 |
BIO19 |
DEM |
Hillshade |
Aspect |
Slope |
|
AdBn01 |
X |
X |
X |
X |
X |
X |
X |
||||||||||||||||
|
AmMn01 |
X |
X |
X |
X |
X |
X |
X |
X |
|||||||||||||||
|
AsAn01 |
X |
X |
X |
X |
X |
X |
|||||||||||||||||
|
AsMa01 |
X |
X |
X |
X |
X |
X |
X |
||||||||||||||||
|
AsMn02 |
X |
X |
X |
X |
X |
X |
X |
X |
X |
||||||||||||||
|
AsSn01 |
X |
X |
X |
X |
X |
X |
|||||||||||||||||
|
BdBn01 |
X |
X |
X |
X |
X |
X |
X |
X |
|||||||||||||||
|
BdPn01 |
X |
X |
X |
X |
X |
X |
|||||||||||||||||
|
BdTc01 |
X |
X |
X |
X |
X |
X |
X |
||||||||||||||||
|
BdTc02 |
X |
X |
X |
X |
X |
X |
X |
X |
X |
||||||||||||||
|
BeBn01 |
X |
X |
X |
X |
X |
X |
X |
X |
X |
X |
|||||||||||||
|
BePn01 |
X |
X |
X |
X |
X |
X |
X |
X |
|||||||||||||||
|
BePn02 |
X |
X |
X |
X |
X |
X |
X |
||||||||||||||||
|
BeTc02 |
X |
X |
X |
X |
X |
X |
X |
||||||||||||||||
|
BmBn01 |
X |
X |
X |
X |
X |
||||||||||||||||||
|
BmBn02 |
X |
X |
X |
X |
X |
X |
X |
X |
X |
||||||||||||||
|
BmPn01 |
X |
X |
X |
X |
X |
X |
|||||||||||||||||
|
BmPn02 |
X |
X |
X |
X |
X |
X |
X |
||||||||||||||||
|
BmTc01 |
X |
X |
X |
X |
X |
X |
X |
||||||||||||||||
|
BsAn02 |
X |
X |
X |
X |
X |
||||||||||||||||||
|
BsAn04 |
X |
X |
X |
X |
X |
X |
X |
||||||||||||||||
|
BsBa02 |
X |
X |
X |
X |
X |
X |
X |
X |
|||||||||||||||
|
BsBa03 |
X |
X |
X |
X |
X |
X |
|||||||||||||||||
|
BsBn02 |
X |
X |
X |
X |
X |
X |
X |
||||||||||||||||
|
BsBn04 |
X |
X |
X |
X |
X |
X |
X |
X |
X |
||||||||||||||
|
BsBn05 |
X |
X |
X |
X |
X |
X |
|||||||||||||||||
|
BsMa01 |
X |
X |
X |
X |
X |
X |
X |
X |
|||||||||||||||
|
BsMa02 |
X |
X |
X |
X |
X |
X |
|||||||||||||||||
|
BsMn02 |
X |
X |
X |
X |
X |
X |
X |
||||||||||||||||
|
BsMn03 |
X |
X |
X |
X |
X |
X |
X |
X |
X |
||||||||||||||
|
BsMn04 |
X |
X |
X |
X |
X |
X |
X |
X |
X |
X |
|||||||||||||
|
BsPa02 |
X |
X |
X |
X |
X |
X |
X |
X |
|||||||||||||||
|
BsPa03 |
X |
X |
X |
X |
X |
X |
X |
||||||||||||||||
|
BsPa04 |
X |
X |
X |
X |
X |
X |
X |
||||||||||||||||
|
BsPn02 |
X |
X |
X |
X |
X |
||||||||||||||||||
|
BsPn04 |
X |
X |
X |
X |
X |
X |
X |
X |
X |
||||||||||||||
|
BsTc05 |
X |
X |
X |
X |
X |
X |
|||||||||||||||||
|
HsSn02 |
X |
X |
X |
X |
X |
X |
X |
||||||||||||||||
|
HsSn04 |
X |
X |
X |
X |
X |
X |
X |
||||||||||||||||
|
HsTc03 |
X |
X |
X |
X |
X |
Fuente: resultados del proceso de modelización. Elaboración propia.
Precipitación actual y futura de los ecosistemas de la Zona Sur.
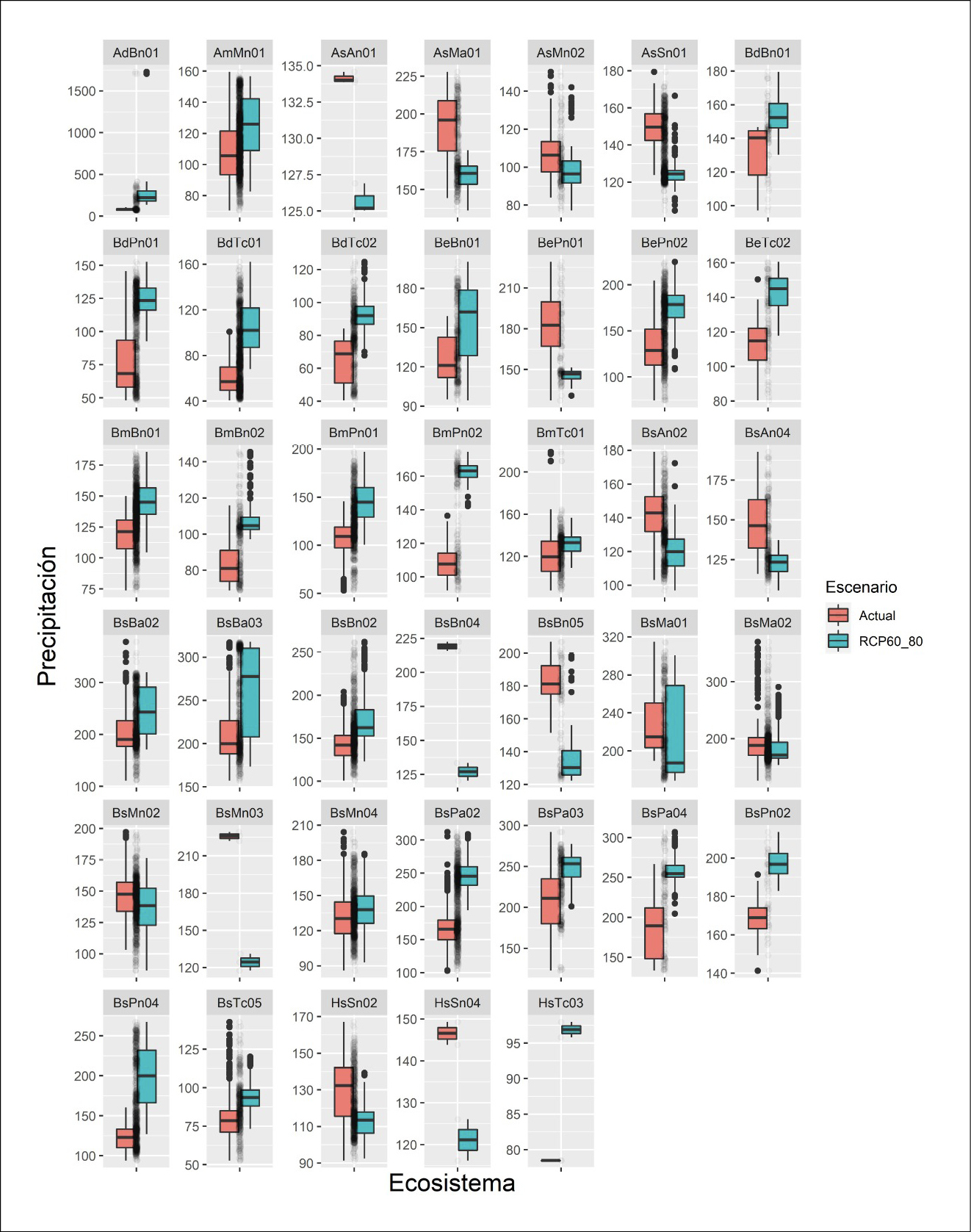
Fuente: resultados del proceso de modelización. Elaboración propia.
Temperatura actual y futura de los ecosistemas de la Zona Sur.

Fuente: resultados del proceso de modelización. Elaboración propia.
